Lysol is a disinfectant designed for use on surfaces and contains phenol, a protoplasmic poison. Poisoning from Lysol can occur through oral ingestion, skin contact, inhalation, and other means. Phenol is mainly metabolized by the kidneys, and after absorption, it causes widespread capillary damage and clotting in superficial blood vessels. This results in urine that is scanty with traces of albumin, and microscopically, blood and calcium oxalate crystals are observed. The urine turns olive green or brown on standing, a phenomenon called carboluria.Studies by J. Jyrkkanen 2020 on antibiotic impacts on mitochondria showed that ROS damage and ruptures of mitochondria resulted in pathology leading to loss of oxidative phosphorylation, likelihood of increase in cancers favorite metabolism aerobic glycolysis, suppression of the p53 DNA repair gene, cardiovascular ailments and immune dysfunction. The similarity of damage reported here by google AI suggest a similar pathology for Lysol type antibiotic cleaners.Otherwise damaged mitochondria induced effects have been observed also for some common pesticides, and mRNA vaccine.The problem arises from the fact that Lysol is an antibiotic that kills bacteria and mitochondria are a highly modified essential symbiotic derivative of an ancient bacteria.
Characteristics
Values
Sensation
Constriction around the throat, burning pain from the mouth to the epigastrium
Taste
Sour and acidic
Urine
Scanty, with traces of albumin
Microscopic Observations
Blood and calcium oxalate crystals
Other Observations
Burns on the face and skin, mucosa of the mouth, tongue, pharynx, and esophagus may be bleached
Phenol Metabolism
Converted into hydroquinone and pyrocatechol, then excreted in urine
Urine Color
Olive green or brown on standing (carboluria)
Impacts on Mitochondria
The active ingredient benzalkonium chloride reveal a pattern of cellular toxicity mediated by mitochondrial damage.
Disruption of Energy Production
QACs which are commonly used in antimicrobial products, have been shown to inhibit mitochondrial respiration specifically at Complex I of the electron transport chain, leading to a decrease in cellular energy (ATP)
Structural Damage or Fragmentation
Exposure to these compounds causes structural disruption, resulting in mitochondrial fragmentation and changes in mitochondrial membrane potential
Induction of Oxidative Stress
Similar to other chemical sanitizers, these compounds can induce the generation of reactive oxygen species (ROS) within the mitochondria, causing further impairment.
Initiation of Apoptosis
Mitochondrial dysfunction caused by these surfactants can trigger the mitochondrial pathway of apoptosis or programmed cell death
Compounds with Documented Mitochondrial Toxicity
Quaternary Ammonium Compounds (QAC’s/BAC’s) induce mitochondrial dysfunction in epithelial cells
Cetylpyridinium Chloride (CPC)CPC causes severe nanostructural disruption, reduces ATP production, and acts as a mitochondrial inhibitor.
Ammonium Hydroxide a caustic compound present in some cleaners can cause severe irritation and potential cellular disruption.
CONCLUSION
These disinfectants can cause significant mitochondrial dysfunction in mammalian cells leading to significant morbidity and mortality. While disinfectant effect may reduce exposure to pathogenic microbes in the short term the harm to beneficial endogenous mitochondria may cause health issues in the long term.
References
Jyrkkanen jccm-aid1104
Jyrkkanen J. Antibiotic induced changes to mitochondria result in potential contributions to carcinogenesis, heart pathologies, other medical conditions and ecosystem risks. J Cardiol Cardiovasc Med. 2020; 5: 163-171
My Lifetime Research Findings:After reading the following and following the links if you want further reading of my literature, try using these search terms after my name: Jorma Jyrkkanen antibiotics, cancer, oncology, cardiology, epigenetics, music, Jormawankenobe, birds, hiking, climate, virology, covid, spike, BWC, geopolitics, WEF, bombing, Deep State, evolution, Darwin, spallation, methylation, bald eagles, Khutzeymateen, Kvenland, extinction, Skagit, Books, blogspot.com, twitter, wordpress.com and follow the links. You may also explore my Bibliography which I add to on occasion. https://jormajyrkkanen.ca/2024/02/14/jorma-antero-jyrkkanens-bibliography-2024-02-14/
3. Ontario Minnesota Pulp and Paper company were liquidating the Ontario boreal forests by fudging regeneration statistics. Chief Forester George Brown shut them down and made them agree to Boreal Forest Clear cut Guidelines and regeneration plans.
4. Vision herbicide, a glyphosate Monsanto product, had 350 ppm of carcinogenic 1,4-dioxane in it and its surfactant polyoxyethyleneamine POEA added to its carcinogenic risk. https://jorma-jyrkkanen.livejournal.com/184872.html
Spallation synthesis of Boron probably made life possible in the universe. It seems we owe our existence to high energy radiation as well as star stuff.
My Conclusion: Cosmic ray spallation is thought to be responsible for the abundance in the universe of some light elements—lithium, beryllium, and boron—as well as the isotope helium-3. They exist only by virtue of their nucleosynthesis during the Big Bang, and also cosmic ray spallation. These lighter elements in the present universe are therefore thought to have been produced through thousands of millions of years of cosmic ray (mostly high-energy proton) mediated breakup of heavier elements, spallation in interstellar gas and dust. This suggests life owes its existence much to spallation by proton radiation collaborating with star stuff.
Pro-vaccination Dr blows whistle on vaccination and finds huge DNA contamination which can increase inflammation and myocarditis, SV40 linked to brain cancer, bone, NHL and malignant mesothelioma and spike protein in heart muscle on autopsy and batch switch from approved to mass produced for public use.
8. I gave evidence at in International Conference on Genetics, Genetic disorders and Stem Cells in Stockholm in 2018 that there is a pandemic of cancers caused by infectious agents ongoing and mouse mammary tumor virus (MMTV), simian virus 40 (SV40) needed to be added to the list of agents and Epstein Barr virus needs special attention. I computed a rough estimate of surplus cancer deaths worldwide attributable to infectious agents without considering gain-of-function bioweapons of approximately 2,365, 920 Deaths in 2023. This is now of course an under-estimate because we need to add the mortality from Turbo cancers from Covid-19 reported by Japanese, UK and American Oncologists. AIDS cancers ex: lung, anal, hepatocellular, Hodgkins lymphoma, oropharyngeal cancers, sars2 covid-19 cancers, SV40 cancers ex: SV40 may contribute to the development of mesotheliomas in humans. Epidemiological studies are needed. SV40, a monkey and now human polyomavirus, was introduced into the human population as a contaminant of vaccines. Merck Virologist Maurice Hilleman bragged about spreading SV40 tumors to Russian Olympians so America would win the Olympics.
I hereby declare that there is a global pandemic of cancers caused by infectious agents, most natural, some with possible increased pathogenicity by gain-of-function and enhanced transmissability (ex: HIV and SV40 by plandemic or injection). There is an increased risk of developing cancer from exposure to these pathogens and also transmit that risk to others by poor hygeine.
Microbiologist Kevin McKernan ran tests on covid vaccine approved and injected and found injected vaccine had DNA contamination and from sequence data was not the one that was approved.
Breaking News Pfizer Lawyer States We did no commit fraud, we delivered the Fraud the Government Ordered. Covid was a DOD fraud all along leading to over 17 Million Killed Worldwide.
Quoting Dr. David Martin: Peter Daszak wrote in 2015 ‘they needed the media to hype up the need for coronavirus vaccines to sell them’. The very next year Dr Ralf Baric wrote in 2016: ‘..’the (WIV) virus was human ready’ a clear indication a weapon was being readies for human transmission. [Seems to me J. Jyrkkanen ‘Collusion and Conspiracy to commit a bioweapon convention violation BWC for profit’]
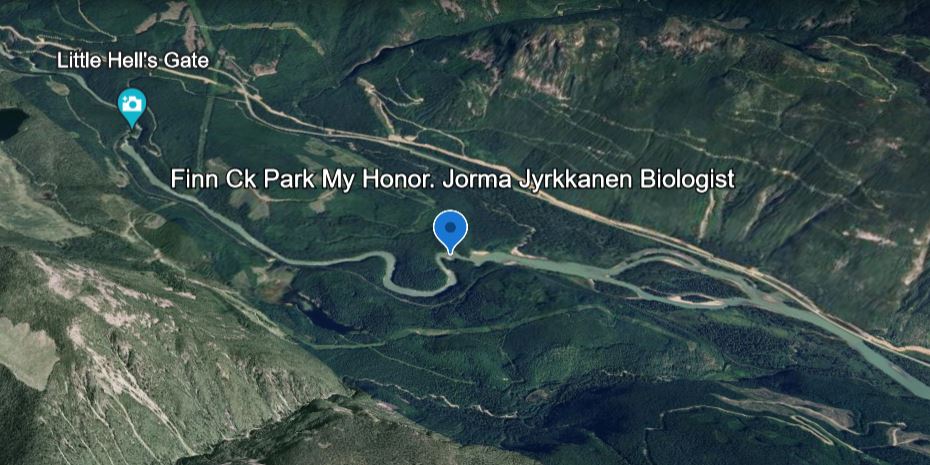
21. How Prince Phillip, The Duke of Edinborough and I deliver the Khutzeymateen Grizzly Sanctuary to the World. I had to put this in because its such a good news story I was involved in.
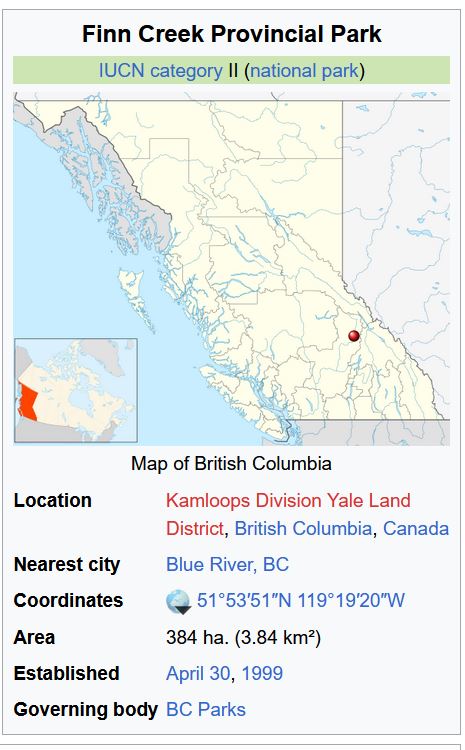
For my conservation and habitat work I was given the honor of having a BC Park Named after me. I asked them to use my nic-name at work “FINN”. Ergo ‘FINN CREEK PARK’.
22. I computed the eigenvectors and principle eigenvalue of Squamish Wintering Bald Eagles to what I think is a close approximation from real data I happily gathered over four years and believe have nailed it. It tells me eagles have evolved a density dependent feedback system of population regulation that leads to growth and a stable age distribution. I Post it because its cool and used my UBC Zoology training beautifully. I think I deserve a PHd for this.
Computing Eigenvalues and Eigenvectors for Squamish River Wintering Bald Eagles. Jorma Jyrkkanen, jjyrkkanen76@outlook.com 3 Dec 2018 Background I fiddled with my 4 years of Squamish Wintering Eagles demographic data and fudged a bit of the model to add males in the Reproduction and got an interesting growth rate, lambda1 and a great Eigenvector or stable age distribution. Here it is. Will play with it until I am happy with data fit and may extend it out to 15-20 years age classes. It’s a dynamic model amenable to gaming based on thousands of field observations.
Discussion What I saw from year to year though were what looked like compensatory density dependent adjustments in age specific survival so it flip flopped p due to competition from yr to yr.
This fact is not amenable to deterministic Markov models like Leslie UNLESS you ‘game it’ or program in ‘if Sxi > or < Sxi critical-then-change-Sxi+ or-n’ ie compensatory survival feedbacks to other age classes. I urge population biology students to try it and watch amazing responses to population and age structure. I used an online Calculator bandicoot.maths.adelaide page written by Mathew Roughan.
Figure 1. Sample Leslie Matrix and Start Population Vector with test age specific reproductive rates for a Female Population based on Hypothetical Statistics to Observe Deterministic Model Responses in Total Population, Age Classes and Growth Rates (Primary Eigenvalue) and proportional age class structure (Eigenvectors).December 3, 2018Created by Jorma A. Jyrkkanen Note. The last three age classes are all adults and there is some presumption of productivity from four year class Bald Eagles ie R4=0.1
Eagles Run 2 with a Working Survivorship Population Vector but discrepancy in observed and model that works.
This latter run has a high crash potential (CP) in 8 yrs and a lot of fluctuation. For a more realistic model I have to incorporate my field data.
The bottom box has the stable age structure eigenvector and age specific eigenvalues. The latter more realistic run works but stability hangs on a few percentage survivorships of juveniles age classes and only includes 8 Age classes total.
Conclusion I concluded that the dynamic nature of feedbacks assures high survival of recruits to replace low adult mortality. Note how with even static Survivorships (Sij diagonals in L) there is an innate generation of wild fluctuations in age-class abundance in the curves at the bottom but I caution that in the field this is difficult to tease apart from random effects of migration linked as it is to weather and food availability. This phenomenon no doubt leads a lot of people to think a population is in trouble when only short-term observations are made and it is not true. Time lags are working to smooth out the age class distributions and real population. You only come to know these statistics as probably true after years of observation. I did 4 with thousands of observations and it was just starting to sink in. Keep in mind that these parameters apply only to the female side of the population. If you add the males it will approximately double.
I was a herd animal til I had a good look around. The Bulls liked the Lambs best.
FUNDING: I only received remuneration on discoveries 1 and 3. The remainder of all my scientific work and the thousands of hours I put in to them has been paid for by me and the hardship it imposed on my family. If some philanthropist has funds to chip in and reimburse me for my work for humanity and nature it would be helpful. My situation is very humble and I could do more with help. To help contact me by text message and we can talk on telegram.
I dedicate my life’s research to my Father and Mother who gave me life. Dad owed his life to a Railway spike. Attacked by Russians he ran out of bullets and a Soviet soldier came to shoot him but dad got him in the heart with a long thrown railway spike, took his firearm and continued the battle which the Finns won. None of my work would have happened without his strength and courage and a railway spike.
Jorma A Jyrkkanen, Bsc, PDP, Researcher. 2023-07-02
My Concerns
It has been found that antibiotic treatment will lead to an increase of reactive oxygen and lipid peroxide, two potential mutagens and carcinogens and this may even rupture the mitochondria with subsequent serious loss of immune and metabolic functionality, for example loss of oxidative phosphorylation, ATP and NAD and increased immune dysfunction leading to cardiovascular pathologies and increased cancers. Chinese researchers also found that a number of common pesticides will also rupture mitochondria. This may lead to similar results. The big surprise was that covid virus may also attacks and destroy the mitochondria. Will the vaccine likewise have deleterious effects on mitochondrial contributions? From these findings it is no surprise that lipid peroxide titre in patients has been found to coincide with severity of the infection. To my astonishment antibiotics were administered to severely ill covid patients and it came as no surprise to me that so many died. Did the antibiotic contribute to their deaths? I suspect that it did.
What needs to happen is body burden and past history of antibiotics use and type of antibiotic need to be assessed in every case, and exposures to common pesticides as well before any administration and lipid peroxide titre needs to be taken into account. The Mitochondria is after all an ancient bacteria and is vulnerable to biocides and this must be taken into consideration. Mitochondria are also absolutely critical for normal cardiovascular and immune function so all care and attention must be taken to ensure no damage occurs to this organelle by any treatment.
Antibiotics Use in Hospitalised COVID-19 Patients in a Tertiary Care Centre: A Descriptive Cross-sectional Study and Heart Mitochondria. JORMA JYRKKANEN 2023-03-23
Thapa B, Pathak SB, Jha N, Sijapati MJ, Shankar PR. Antibiotics Use in Hospitalised COVID-19 Patients in a Tertiary Care Centre: A Descriptive Cross-sectional Study. JNMA J Nepal Med Assoc. 2022 Jul 1;60(251):625-630. doi: 10.31729/jnma.7394. PMID: 36705203; PMCID: PMC9297358.
ABSTRACT Introduction:
Antimicrobial resistance is a global health problem. The widespread and improper antibiotics use is the leading cause of antimicrobial resistance. Bacterial co-infection in COVID-19 patients is the basis for the use of antibiotics in the management of COVID-19. COVID-19 pandemic has seriously impacted antibiotic stewardship and increased the global usage of antibiotics, worsening the antimicrobial resistance problem. The use of antibiotics among COVID-19 patients is high but there are limited studies in the context of Nepal. This study aimed to find out the prevalence of antibiotic use among hospitalised COVID-19 patients in a tertiary care centre.
Introduction:
Antimicrobial resistance is a global health problem. The widespread and improper antibiotics use is the leading cause of antimicrobial resistance. Bacterial co-infection in COVID-19 patients is the basis for the use of antibiotics in the management of COVID-19. COVID-19 pandemic has seriously impacted antibiotic stewardship and increased the global usage of antibiotics, worsening the antimicrobial resistance problem. The use of antibiotics among COVID-19 patients is high but there are limited studies in the context of Nepal. This study aimed to find out the prevalence of antibiotic use among hospitalised COVID-19 patients in a tertiary care centre. Methods:
A descriptive cross-sectional study was conducted on hospitalised COVID-19 patients from April 2021 to June 2021 in a tertiary care centre. Ethical approval was taken from the Institutional Review Committee (Reference number: 2078/79/05). The hospital data were collected in the proforma by reviewing the patient’s medical records during the study period of 2 months. Convenience sampling was used. Point estimate and 95% Confidence Interval were calculated. Results:
Among 106 hospitalised COVID-19 patients, the prevalence of antibiotics use was 104 (98.11%) (95.52-100, 95% Confidence Interval). About 74 (71.15%) of patients received multiple antibiotics. The most common classes of antibiotics used were cephalosporins, seen in 85 (81.73%) and macrolides, seen in 57 (54.81%) patients. Conclusions:
The prevalence of antibiotics use among hospitalised COVID-19 patients was found to be higher when compared to other studies conducted in similar settings. Keywords: antibiotics, bacterial infection, co-infection, COVID-19 Go to: INTRODUCTION
Antimicrobial resistance (AMR) is a major threat to global public health due to the increasing incidence of resistant human pathogens.1,2 The widespread and improper use of antibiotics is the leading cause of AMR.1 Coronavirus Disease 2019 (COVID-19) is a viral disease thus untreatable by antibiotics, but the viral respiratory infections may clinically progress to bacterial pneumonia requiring antibiotic administration.2 This co-pathogenesis is the basis for use of antibiotics in COVID-19. But appropriate use of antibiotics is utmost to prevent AMR.
COVID-19 pandemic has seriously impacted antibiotic stewardship and single-handedly increased the global usage of antibiotics, causing a cascading effect on the AMR problem. The use of antibiotics among COVID-19 patients is high but there are limited studies in the context of Nepal.3-5
This study aimed to find out the prevalence of antibiotics use among COVID-19 patients of a tertiary care centre. Go to: METHODS
A descriptive cross-sectional study was conducted at KIST Medical College and Teaching Hospital after taking ethical approval from the Institutional Review Committee (Reference number: 2078/79/05). The study was conducted during the study period from 6 August 2021 to 6 October 2021 during which hospitalised COVID-19 patients admitted from April 2021 to June 2021 were studied. All the COVID-19 cases confirmed by reverse transcriptase polymerase chain reaction (RT-PCR) test who were admitted in the dedicated COVID-19 ward, high dependency unit (HDU) and intensive care units (ICU) were enrolled. Patients who had incomplete documentation were excluded from the study. Convenience sampling was used. The sample size was calculated using the following formula: n=Z2×p×qe2=1.962×0.50×0.500.102=97
Where,
n = minimum required sample size
Z = 1.96 at 95% Confidence Interval (CI)
p = prevalence taken as 50% for maximum sample size calculation
q = 1-p
e = margin of error, 10%
Minimum sample size calculated was 97. However, we enrolled 106 cases. The collected data from hospital records was entered in the proforma by reviewing the patient’s medical records during the study period of two months. Demographic profile of patients like age and sex, clinical profile like co-morbidity and disease severity, management profile like level of care required for patients’ treatment, number and type of antibiotics used, route of antibiotic administration, duration of antibiotics used, and estimated cost of antibiotics used for the treatment were assessed. The patients who were treated with at least one antibiotic were included. All the cases were classified as a mild disease, moderate disease, or severe disease.6
In our study, 17 different antibiotics were used belonging to seven different antibiotic classes. They are namely cephalosporin (ceftriaxone, cefixime, cefoperazone, and cefepime), macrolides (azithromycin, clindamycin, and erythromycin), penicillin group (piperacillin, amoxicillin), quinolones (moxifloxacin, levofloxacin, ciprofloxacin), imidazoles (metronidazole), carbapenem (meropenem) and beta-lactamase inhibitors (clavulanic acid, tazobactam, sulbactam).
The data were entered and analysed using IBM SPSS Statistics 21.0. Point estimate and 95% CI were calculated. Go to: RESULTS
Among 106 hospitalised COVID-19 patients, the prevalence of use of antibiotics was 104 (98.11%) (95.52-100, 95% CI). The mean number of antibiotics used per patient was 1.86 ±0.64. A total of 74 (71.15%) patients were under two or more antibiotic therapy. Around 60 (57.69%) patients were treated with intravenous as well as per oral route of administration of antibiotics.
The mean number of days of admission was 6.44±4.81 days. The mean duration of antibiotics use was 6.33 ±2.72 days. About 30 (28.85%) received a 5 day course of antibiotics while 25 (24.04%) patients received a 7 days course of antibiotics. Only 23 (22.12%) received antibiotic therapy for more than 7 days. The mean estimated expenditure on antibiotics was NPR 4,645±8, 498 (USD 38.71±70.82) (Table 1). Table 1 Use of antibiotics in management of COVID-19 patients (n = 104). Characteristics n (%) Number of antibiotics used 1 30 (28.85) 2 59 (56.73) 3 15 (14.42) Route of antibiotics Intravenous route only 37 (35.58) Per oral route only 7 (6.73) Intravenous and per oral route 60 (57.69) Total number of days of antibiotics use ≤7 days 81 (77.88)
7 days 23 (22.12) Estimated expenditure on antibiotics therapy NPR (US dollar) ≤1,200 (USD 10) 43 (41.35) 1201 to 6,000 (USD 11 to 50) 42 (40.38) 6,001 to 12, 000 (USD 51 to 100) 8 (7.69) 12,001 to 24,000 (USD 101 to 200) 7 (6.73) 24,000 (USD 200) 4 (3.85) Open in a separate window
The mean age of the patients was 55.84±18 years. A total of 54 (51.92%) patients were males. Around 59 (56.73%) had at least one comorbid condition with the most common conditions being hypertension seen in 39 (37.50%) and diabetes mellitus seen in 23 (22.16%) (Table 2). Table 2 Demographic characteristics of hospitalised COVID-19 patients who received antibiotic therapy (n= 104). Age group n (%) ≤20 years 5 (4.81) 21 to 40 years 18 (17.31) 41 to 60 years 37 (35.58) 61 to 80 years 37 (35.58)
80 years 7 (6.73) Sex Males 54 (51.92) Females 50 (48.08) Comorbidities Diabetes mellitus 23 (22.16) Hypertension 39 (37.50) Chronic Obstructive Pulmonary 10 (9.62) Disease (COPD) Hypothyroidism 12 (11.54) Psychiatric illness 2 (1.92) Heart failure 2 (1.92) Autoimmune disease 2 (1.92) Chronic kidney disease 1 (0.96) Open in a separate window
Severe COVID-19 represented 37 (35.58%) of total patients, 16 (15.38%) of them were managed in ICU with ventilator support. Moderate COVID-19 cases also accounted for 37 (35.58%) of total patients. These patients were mostly managed in a dedicated COVID-19 ward with 2 (1.92%) cases managed in ICU and 2 (1.92%) in HDU. All 30 (28.84%) mild cases were managed in the ward (Table 3). Table 3 COVID-19 severity and level of care received by the patients who received antibiotic therapy (n = 104). Level of care Mild n (%) Moderate n (%) Severe n (%) Total n (%) ICU with ventilator support – – 16 (15.38) 16 (15.38) ICU without ventilator support – 2 (1.92) 4 (3.85) 6 (5.77) HDU – 2 (1.92) 15 (14.42) 17 (16.35) Ward 30 (28.84) 33 (31.73) 2 (1.92) 65 (62.50) Open in a separate window
The most common class of antibiotics used was cephalosporins in 85 (81.73%) patients followed by macrolides in 57 (54.81 %). Cefixime used in all cases was a substitute for ceftriaxone in oral form in 18 (17.31%) patients who were previously prescribed ceftriaxone. Beta-lactamase inhibitors were used in 35 (33.65%) in conjunction with penicillin (amoxicillin) or cephalosporin group of drugs (cefoperazone, cefepime). The most common combination used was cephalosporin with macrolides at 38 (36.54%) (Table 4). Table 4 Types of antibiotics used in management of COVID-19 patients (n = 104). Antibiotics n (%) Cephalosporins prescribed parenterally 85 (81.73) Ceftriaxone 76 (73.08) Cefepime sulbactam 3 (2.88) Cefoperazone sulbactam 6 (5.77) Cephalosporins prescribed enterally 18 (17.31) Cefixime 18 (17.31) Macrolides 57 (54.81) Azithromycin 55 (52.88) Erythromycin 1 (0.96) Clindamycin 1 (0.96) Penicillins 26 (25.00) Piperacillin tazobactam 14 (13.46) Amoxicillin clavulanic acid 12 (11.54) Quinolones 11 (10.58) Moxifloxacin 9 (8.65) Levofloxacin 2 (1.92) Ciprofloxacin 1 (0.96) Imidazoles 10 (9.62) Metronidazole 10 (9.62) Carbapenems 4 (3.85) Meropenem 4 (3.85) THIS IS A COCKTAIL THAT IS VERY DANGEROUS TO HEART MITOCHONDRIA-JORMA JYRKKANEN COMMENT DISCUSSION
The prevalence of use of antibiotics was 98.1%. About 71.15% patients were treated with two or more antibiotics. The mean number of antibiotics used per patient was 1.86. The mean duration of antibiotics use was 6.33 days. Seventeen different antibiotics were used belonging to 7 different antibiotic classes. The most common class of antibiotics used was cephalosporin at 85 (81.73%) and macrolides at 57 (54.81%).
Even before the COVID-19 pandemic, AMR was projected to become responsible for approximately 10 million deaths worldwide in the coming three decades.7 COVID-19 has undoubtedly affected antibiotic stewardship and has increased antibiotic consumption patterns globally, adding to the already existing global AMR problem. Because of this, the mortality due to AMR is expected to be higher in post COVID era.2 This pandemic has disrupted health delivery systems worldwide. This has increased the overuse of antibiotics, eventually leading to resistant organisms requiring aggressive treatment.8 Thus AMR is a problem of greater concern than COVID-19 which has unfortunately been overshadowed amidst the pandemic.7,9 Increased use of antibiotics is more challenging, especially in the low and middle-income countries (LMIC) due to the inefficiency and inadequacy of health care services.2
The Infectious Diseases Society of America (IDSA) states that only 8% of the COVID-19 patients acquired bacterial/fungal superinfections requiring antibiotics.10 However, a study showed 72% of COVID-19 patients received empirical broad-spectrum antibiotics, even when bacterial coinfection was absent.11 Current World Health Organization guidelines indicate that antibiotics should not be prescribed in mild or moderate COVID-19 cases unless there are pre-existing symptoms of bacterial co-infection. Furthermore, when treating severe cases with an empirical antimicrobial agent, the overall condition of the patient, local bacterial epidemiology, and clinical judgement should be integrated, to ensure judicial antimicrobial usage.12 In COVID-19 patients, antibiotics are used for potential anti-inflammatory, immune-modulating, and potential antiviral properties. But the antiviral mechanism of these agents is doubtful. This widespread antibiotic use is likely to worsen preexisting AMR crisis.13
The influenza pandemic was largely a problem of viral infection complicated by bacterial co-pathogenesis.14 This has been our basis for use of a wide range antibiotics empirically though COVID-19 is primarily a viral pathology and is not conventionally treated with antibiotics.
In a study, 71.00% of the hospitalised COVID-19 patients received antibiotics despite a confirmed bacterial co-infection rate of only 1%.3 Antibiotic was used in 95.00% COVID-19 patients when secondary bacterial infection was only found in 15.00%.4 A systematic review showed the mean rate of antibiotic use was 74.00 %.5 In our study, the prevalence of use of antibiotics was 98.10% which is very high when compared to above studies. In most cases antibiotics use often empirical. Empiric antibiotics were often used for the concern of community-acquired pneumonia (89.00%).15 This showed that antibiotic therapy has been used often empirically in the majority of patients even when very few were proven to have bacterial coinfection.
In our study, 17 different antibiotics belonging to seven antibiotic classes were used. Similar to our study a wide range of antibiotics use was documented in other studies.1,5,10,13,15-18 Many other classes of antibiotics other than above were used in other studies for the management of COVID-19 patients. They are aminoglycosides,1 glycopeptide antibiotic like vancomycin and teicoplanin,10 oxazolidinones like linezolid, tetracycline and cyclic lipopeptides like daptomycin.17 Most of these are newer classes of antibiotics and increased use of these should raise a red flag among concerned clinicians, pharmacists, microbiologists, public health experts, hospitals, local authorities as well as regulatory bodies.
Carbapenem, fluoroquinolones, and aminoglycoside were highly prevalent in ICU patients.1 Similar to this carbapenem was exclusively used for ICU patients in our study. Other commonly used antibiotics among ICU patients were fluoroquinolones, cephalosporin, piperacillin with tazobactam, and macrolides. In general, ICU addmission compromises of a very sick patient with superadded bacterial infection and in regard to COVID-19, it comprises of severe COVID-19 infection often requiring ventilatory support. In such conditions, it is common practice to use multiple higher and broad-spectrum antibiotics.
The common antibiotics in use were ceftriaxone (54.00%), vancomycin (48.00%), azithromycin (47.00%), and cefepime (45.00%).10 In our study ceftriaxone (73.08%) and azithromycin (52.88%) were widely used but cefepime was used in 2.88% of patients and vancomycin was not used at all. Higher antibiotics like cefepime and vancomycin should only be used when there is a valid indication, otherwise, it may result in resistant infection which will be very hard to treat.
Ceftriaxone and azithromycin are often the most common antibiotics used in the management of COVID-19 patients.10,13,15,16 Most of the local guidelines as well as some international guidelines advocate for use of these antibiotics based on the epidemiology of local pathogens and resistance patterns. The advantage of the use of these antibiotics is that it covers most of the opportunistic pathogens that could cause secondary infection in COVID-19. But on the other hand, wide and inappropriate use of these antibiotics can lead to with emergence resistance of these common, cheap, and very efficient antibiotics.
Macrolide, specifically azithromycin, was the most common antibiotic used in the clinical management of COVID-19.13 Macrolides, particularly azithromycin, were used in the treatment of more than half of the patients in our study. These drugs are often used to cover atypical organism that have the potential to cause a secondary infection.10,11
Fluoroquinolones were most used, (56.80%), followed by ceftriaxone (39.50%), then azithromycin (29.10%), and carbapenems were only used in two patients.18 Unlike to above study, in our study fluoroquinolones were used less (10.58%) and ceftriaxone and azithromycin was basically used in most patients. But similarity was observed between ours and the above study regarding the use of carbapenems, which was used in 3.85% of patients. Wide use of carbapenem, used in up to 40.10% of patients was also reported.5 carbapenem is often used as a reserved antibiotic for severe infection thus minimal use of these in COVID-19 signifies the presence of good antibiotics stewardship and antibiotic management system among concerned institutions while wide use can signify the opposite.
The most common antibiotic used was third-generation cephalosporin (ceftriaxone) (53.80%), moxifloxacin (29.50%), and doxycycline (25.40%).5 Similar to the above study the most common class of antibiotics used in our study was cephalosporin (81.74%), around 90.00% of which was third-generation cephalosporin namely ceftriaxone (76/85). But unlike the above, Moxifloxacin was used in only 8.65% and doxycycline or other tetracycline group of drugs was not used at all in our study.
A study showed all patients were receiving at least one antibiotic with 31.08% receiving a single antibiotic and 68.91% receiving multiple antibiotics.5 Similar to this study, 98.10% of patients in our study received at least one antibiotic, 28.35% received single antibiotic agents and 71.15% received multiple antibiotics. But the mean number of antibiotics used in the study above was 2.02 which is more when compared to 1.85 in our study. In 34.6%, three antibiotics were given simultaneously while 9.6% received only one antibiotic.16 Unlike the above, in our study three antibiotics were used in only 14.42% and a single antibiotic was used in 28.85%. The use of multiple antibiotics is worrisome as this might represent unchecked use of antibiotics which will contribute to the worldwide problem of AMR.
In our study, patients with comorbidity were found to have received multiple antibiotics (72.90%). Around 74.00% of patients with diabetes were on multiple antibiotics. Similar findings were documented in another study.5 This might be due to COVID-19 patients with comorbidities like diabetes, airway diseases, hypertension being at greater risk of developing secondary bacterial infection.19 After predicting the risk of secondary infection, multiple antibiotics were used empirically in those patients.
Overall, patients presenting with severe disease received more antibiotics.5 This is also true for our study. COVID-19 patients with co-morbidities and severe COVID-19 are the two most vulnerable groups of patients, thus multiple antibiotics have been found to be used liberally in these patients. These situations can be dealt with systematically by establishing standard antibiotic prescribing guidelines considering local pathogens and sensitivity patterns to antibiotics. This could be further reinforced by appropriate clinical knowledge, laboratory facilities, and surveillance systems.
In our study the mean duration of antibiotics treatment was 6.33 days which is nearly half when compared to 12.71 days with a range from 3 days to 23 days.16 Similar result was found in another study.18 Both of these, decreased and extended duration of antibiotic treatment might represent inappropriate and improper use of antibiotics regimen. Because both, underuse and overuse of antibiotics can result in the emergence of resistance.
Our study is a single centred study and has a small sample size. Therefore, our findings may not be generalizable to other settings. Go to: CONCLUSIONS
The prevalence of use of antibiotics among hospitalised COVID-19 patients was found to be higher when compared to other studies conducted in similar settings. Potential bacterial co-infection has been the basis for the use of antibiotics in the management of COVID-19 patients. The rate and number of antibiotics used for mild to moderate disease were also high. The common class of antibiotics used are cephalosporin and macrolides namely ceftriaxone and azithromycin. Higher class antibiotics were mostly used in the management of severe disease in ICU and with ventilator support. However, judicial use of antibiotics among COVID-19 patients with variable severity especially among those admitted in ICU and on ventilatory support could be promoted in order to reduce AMR during this COVID-19 pandemic. Robust antibiotic stewardship programs and surveillance systems should be implemented. Go to: ACKNOWLEDGMENTS
The authors would like to acknowledge KIST Medical College and Teaching Hospital for their support. Go to: Conflict of Interest
None. Go to: REFERENCES
Zeshan B, Karobari MI, Afzal N, Siddiq A, Basha S, Basheer SN, et al. The Usage of Antibiotics by COVID-19 Patients with Comorbidities: The Risk of Increased Antimicrobial Resistance. Antibiotics (Basel). 2021 Dec 29;11(1):35. doi: 10.3390/antibiotics11010035. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Rizvi SG, Ahammad SZ. COVID-19 and antimicrobial resistance: A cross-study. Sci Total Environ. 2022 Feb 10;807(Pt 2):150873. doi: 10.1016/j.scitotenv.2021.150873. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Chen N, Zhou M, Dong X, Qu J, Gong F, Han Y, et al. Epidemiological and clinical characteristics of 99 cases of 2019 novel coronavirus pneumonia in Wuhan, China: a descriptive study. Lancet. 2020 Feb 15;395(10223):507–13. doi: 10.1016/S0140-6736(20)30211-7. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Zhou F, Yu T, Du R, Fan G, Liu Y, Liu Z, et al. Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: a retrospective cohort study. Lancet. 2020 Mar 28;395(10229):1054–62. doi: 10.1016/S0140-6736(20)30566-3. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Molla MMA, Yeasmin M, Islam MK, Sharif MM, Amin MR, Nafisa T, et al. Antibiotic prescribing patterns at COVID-19 dedicated wards in Bangladesh: findings from a single center study. Infect Prev Pract. 2021 Jun;3(2):100134. doi: 10.1016/j.infpip.2021.100134. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Indian Council of Medical Research. Clinical guidance for management of adult COVID-19 patients [Internet]. New Delhi (IN): Indian Council of Medical Research; 2022. Jan 14, [2022 Jan 14; ]. [2022 Feb 20; ]. https://www.icmr.gov.in/ctechdocad.html Available from: [Google Scholar]
Barocas JA, Savinkina A, Lodi S, Epstein RL, Bouton TC, Sperring H, et al. Projected long-term impact of the COVID-19 pandemic on hepatitis C outcomes in the United States: a modelling study. Clin Infect Dis. 2021 Sep 9;:ciab779. doi: 10.1093/cid/ciab779. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Lucien MAB, Canarie MF, Kilgore PE, Jean-Denis G, Fenelon N, Pierre M, et al. Antibiotics and antimicrobial resistance in the COVID-19 era: Perspective from resource-limited settings. Int J Infect Dis. 2021 Mar;104:250–4. doi: 10.1016/j.ijid.2020.12.087. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Neto AGM, Lo KB, Wattoo A, Salacup G, Pelayo J, DeJoy R, et al. Bacterial infections and patterns of antibiotic use in patients with COVID-19. J Med Virol. 2021 Mar;93(3):1489–95. doi: 10.1002/jmv.26441. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Rawson TM, Moore LSP, Zhu N, Ranganathan N, Skolimowska K, Gilchrist M, et al. Bacterial and fungal coinfection in individuals with Coronavirus: A rapid review to support COVID-19 antimicrobial prescribing. Clin Infect Dis. 2020 Dec 3;71(9):2459–68. doi: 10.1093/cid/ciaa530. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Varghese GM, John R, Manesh A, Karthik R, Abraham OC. Clinical management of COVID-19. Indian J Med Res. 2020 May;151(5):401–10. doi: 10.4103/ijmr.IJMR_957_20. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Yacouba A, Olowo-Okere A, Yunusa I. Repurposing of antibiotics for clinical management of COVID-19: a narrative review. Ann Clin Microbiol Antimicrob. 2021 May 21;20(1):37. doi: 10.1186/s12941-021-00444-9. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Morens DM, Taubenberger JK, Fauci AS. Predominant role of bacterial pneumonia as a cause of death in pandemic influenza: implications for pandemic influenza preparedness. J Infect Dis. 2008 Oct 1;198(7):962–70. doi: 10.1086/591708. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Wei W, Ortwine JK, Mang NS, Joseph C, Hall BC, Prokesch BC. Limited Role for Antibiotics in COVID-19: Scarce evidence of bacterial coinfection. [2022 Feb 20; ];medRxiv [Preprint]. 2020 Jun 16; doi: 10.1101/2020.06.16.20133181. https://www.medrxiv.org/content/10.1101/2020.06.16.20133181v1 Available from: [CrossRef] [Google Scholar]
Mustafa L, Tolaj I, Baftiu N, Fejza H. Use of antibiotics in COVID-19 ICU patients. J Infect Dev Ctries. 2021 Apr 30;15(4):501–5. doi: 10.3855/jidc.14404. [PubMed] [CrossRef] [Google Scholar]
Grau S, Echeverria-Esnal D, Gomez-Zorrilla S, Navarrete-Rouco ME, Masclans JR, Espona M, et al. Evolution of antimicrobial consumption during the first wave of COVID-19 pandemic. Antibiotics (Basel). 2021 Jan 29;10(2):132. doi: 10.3390/antibiotics10020132. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Zhu N, Zhang D, Wang W, Li X, Yang B, Song J, et al. A Novel Coronavirus from patients with pneumonia in China, 2019. N Engl J Med. 2020 Feb 20;382(8):727–33. doi: 10.1056/NEJMoa2001017. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Morgan DJ, Casulli J, Chew C, Connolly E, Lui S, Brand OJ, Rahman R, Jagger C, Hussell T. innate immune cell suppression and the link with secondary lung bacterial pneumonia. Front Immunol. 2018 Dec 14;9:2943. doi: 10.3389/fimmu.2018.02943. [PMC free article] [PubMed] [CrossRef] [Google Scholar] THE PROBLEM ARISES FROM THE IMPACT OF A HEAVY DOSE OF ANTIBIOTICS ON THE MITOCHONDRIA AND ALSO THE LOSS OF IMMUNE SYSTEM FUNCTION COINCIDENT WITH LOSS OF MITOCHONDRIAL FUNCTION
Jyrkkanen, Jorma. (2020). Antibiotic induced changes to mitochondria result in potential contributions to carcinogenesis, heart pathologies, other medical conditions and ecosystem risks. Journal of Cardiology and Cardiovascular Medicine. 5. 163-171. 10.29328/journal.jccm.1001104.
In addition to providing energy to support the synthesis of the macromolecules essential for immune cell proliferation, mitochondria also act as signaling organelles, driving activation of immune cells via metabolic intermediates, mitochondrial DNA (mtDNA), and reactive oxygen species (ROS).Mar 22, 2022 Introduction
Immune dysregulation, characterized by an imbalance between a systemic inflammatory response syndrome and a compensatory anti-inflammatory response syndrome, is often observed in critically ill patients [1, 2]. This imbalance between the pro- and anti-inflammatory responses frequently leads to immunoparalysis in critically ill patients, rendering them more susceptible to further infections, and is associated with increased mortality [3]. Currently, no effective treatments are available to restore immune homeostasis and reduce mortality in these patients, largely due to the heterogeneity in patients’ immune status and more importantly the lack of understanding of the underlying cause of such immune dysfunction [2, 4]. Immune response is not a standalone process but is interconnected with other cellular activities, a very important one of which is cellular metabolism. Metabolic pathways and immune response are tightly intertwined both in health and in disease [5]. The link between immune cell function and mitochondrial function is now well recognized and a field known as “immunometabolism” is dedicated to understanding the relationship between immune and metabolic pathways [6,7,8]. Mitochondria play a crucial role in regulating not only the growth, but also the function, of immune cells. In addition to providing energy to support the synthesis of the macromolecules essential for immune cell proliferation, mitochondria also act as signaling organelles, driving activation of immune cells via metabolic intermediates, mitochondrial DNA (mtDNA), and reactive oxygen species (ROS). In addition, mitochondrial dynamics (fusion and fission), biogenesis (synthesis of new mitochondria), and mitophagy (degradation of damaged mitochondria) also play important roles in regulating immune cell functions. Knowledge in immunometabolism in critical illness, in particularly sepsis, opens up a new paradigm in patient care. Potential therapies targeting metabolic pathways, instead of solely immune-related pathways, might be the way to repair cellular function and restore immune homeostasis [4]. The other aspect of immunometabolism—looking at how immune responses influence metabolic pathways—is equally important, but beyond the scope of this review. Interaction between metabolism and immune response at the organ level has been reviewed elsewhere [6]. Mitochondrial Machinery That Mediates and Regulates Immune Responses in Critical Illness
Apart from being the powerhouse of the cell, the mitochondrion has emerged as a signaling hub that shapes and modulates how the immune system responds to infection or trauma. Mitochondrial dysfunction is evident in leukocytes from critically ill patients, and is believed to be the underlying cause of immunoparalysis and may account for the development of organ dysfunction [7,8,9]. Early recovery of mitochondrial function correlates with improved recovery in critically ill patients [10]. Metabolic Reprogramming
The immune-regulating mitochondrial machinery is a complex network involving many pathways and mechanisms that diverge and converge at various levels. Metabolic reprogramming is one mechanism that has been well studied in both innate and adaptive immune cells. Immune cells at different activation states (quiescent vs. activated), or with different functions (pro-inflammatory vs. anti-inflammatory), and different cell types (granulocytes, macrophages, dendritic cells, T- and B-lymphocytes), make use of different metabolic pathways (e.g., glycolysis, oxidative phosphorylation, fatty acid metabolism) to produce ATP [11]. The choice of different metabolic pathways, supports the energy demand of cells at different activation state. For example, upon infection or stimulation, immune cells become activated and produce cytokines and hence tend to favor glycolysis over oxidative phosphorylation for fast turnaround of ATP. Although the same amount of starting material, such as glucose, is used, oxidative phosphorylation generates 18 times more ATP than glycolysis, although is a lot slower. On the other hand, the choice of metabolic pathway determines the fate of the immune cells, i.e., naïve or memory, effector or regulatory, etc. However, the environment that the cells are in in the first place, triggers the changes in the metabolic pathways. The overall trend is that neutrophils, inflammatory macrophages (M1 macrophages), activated effector T cells, and dendritic cells rely more on aerobic glycolysis, whereas alternatively polarized macrophages (M2 macrophages), regulatory T cells (Tregs), and memory T cells prefer oxidative phosphorylation and fatty acid oxidation for energy production [8, 11, 12]. Metabolic reprogramming serves an important role in catering for the immune cells’ energy demand at different phases of their activation and proliferation. However, imbalance across the metabolic pathways could have serious pathological impact. One example may be the hyperlactatemia often seen in critically ill patients. Increased aerobic glycolysis in the activated immune cells during the initial hyper- inflammatory response is believed to contribute to the increase in blood lactate levels in sepsis [13, 14]. Mitochondrial ROS and mtDNA
Metabolic reprogramming sets the scene for the immune response, which is then subjected to many more modifications and regulations by factors that are directly or indirectly related to mitochondrial metabolism. Two important mitochondria-related immune regulators that have been well studied are mitochondrial ROS and mtDNA. Mitochondrial ROS are produced in healthy mitochondria, as a by-product of oxidative phosphorylation. At low dose, mitochondrial ROS serve important signaling functions, especially in the innate immune response. They are known to mediate NLRP3 inflammasome activation, leading to production of the pro-inflammatory cytokines, interleukin (IL)-1β and IL-18 [8, 15]. Mitochondrial ROS also induce a type-I interferon (IFN) response via mitochondrial antiviral-signaling (MAVS) and the IFN regulatory factor 3 (IRF3) pathway [16]. However, the level of mitochondrial ROS needs to be tightly regulated by the antioxidant system. Excessive mitochondrial ROS can cause oxidative damage to proteins/enzymes involved in oxidative phosphorylation and create mutations in mtDNA, contributing to the immune dysregulations as seen in critical illness [17]. Like mitochon-drial ROS, mtDNA also plays an important role in innate immunity [12]. In healthy cells, mtDNA is located in the matrix of mitochondria, encoding 13 proteins, all of which are components of oxidative phosphorylation. mtDNA is released to the cytosol upon mitochondrial dysfunction which involves changes to the integrity or permeability of the mitochondrial membrane. mtDNA, released into the cytosol, can activate the NLRP3 inflammasome with release of IL-1β and IL-18. Due to its bacterial origin, cytosolic mtDNA also serves as a damage-associated molecular pattern (DAMP), which can be recognized by intracellular pattern recognition receptors (PRRs), such as Toll-like receptor 9 (TLR9), and initiate the nuclear factor-kappa B (NF-κB)-dependent pro-inflammatory signaling pathway. In addition, cytosolic mtDNA can also be sensed by cyclic GMP-AMP synthase (cGAS) and activate the cGAS/stimulator of IFN genes (cGAS/STING) pathway and its downstream IFN response [18]. mtDNA can also be released into the circulation and cause systemic inflammation. Circulating mtDNA has been associated with mortality in critically ill patients [19]. Succinate and Itaconate
In addition to mitochondrial ROS and mtDNA, metabolites such as succinate and itaconate have also emerged as part of immune-regulating mitochondrial machinery [4, 20]. Both succinate and itaconate are intermediates from the tricarboxylic acid (TCA) cycle with opposite effects on the immune response. The TCA cycle generates nicotinamide adenine dinucleotide (NADH) and flavin adenine dinucleotide (FADH2), providing electrons to fuel oxidative phosphorylation. Succinate accumulation occurs under conditions such as hypoxia or inflammation. It can be released from mitochondria into the cytosol and functions as a signal transducer promoting pro-inflammatory gene expression via hypoxia-inducible factor 1α (HIF-1α) activation. Accumulation and oxidation of succinate by succinate dehydrogenase (SDH) in the mitochondria also leads to increased production of mitochondrial ROS via a process called reverse electron transport. This further enhances the pro- inflammatory effect of succinate. Like ROS, the level of succinate needs to be carefully regulated due to its inflammation aggravating effect. Plasma succinate has been proposed as a predictor of mortality for critically ill patients who are severely injured [21].
Itaconate, which is derived from cis-aconitate of the TCA cycle, is a succinate-regulating factor. It is shown to counteract the pro-inflammatory effect of succinate by inhibiting SDH. Itaconate can also be released into the cytosol and activate transcription factor NF-E2 p45-related factor 2 (Nrf2), a master regulator of antioxidant and anti-inflammatory responses [22]. Recently, itaconate has also been shown to inhibit the inflammatory response in macrophages through activating transcription factor 3 (ATF3). Mitochondrial Dynamics
The above mentioned immune-regulating mitochondrial factors are centered around the biochemical aspect of mitochondrial biology. Another important aspect of immune-regulating mitochondrial machinery is mitochondrial dynamics, which is to maintain and provide infrastructural support for the immune response. The size and shape of mitochondria undergo constant change through fusion and fission, which is important for maintaining the health and function of mitochondria. First, fusion incorporates newly synthesized mitochondria (from mitochondrial biogenesis) into the current mitochondrial network. Second, fusion also allows for mixing of proteins and/or mtDNA between the existing mitochondria, which on one hand enhances the metabolic capacity of the mitochondria, and on the other enables the damaged proteins and/or mutated mtDNA to be segregated from the healthy ones. Finally, segregation is achieved via fission and the damaged mitochondria can be destroyed through a process known as mitophagy. The proportion of mitochondria with damaged proteins or mutated mtDNA is kept below a critical threshold level through this process to maintain mitochondrial function [23, 24]. In addition to quality control, mitochondrial fusion and fission also participate in immune regulation. In activated T cells, there is an increase in fission, which creates round and fragmented mitochondria with loose cristae, favoring aerobic glycolysis. And in 146 memory T cells, increased fusion generates elongated mitochondria which favors oxidative phosphorylation and fatty acid oxidation [8, 25]. Immunometabolism: The Perfect World Scenario vs. the Critical Illness Scenario
So far, we have presented a list of mitochondrial components that are thought to play important roles in regulating the immune response. Our list is far from complete, but does highlight a few mechanisms that could relate to the development of immune dysregulation in critical illness. Figure 1 illustrates what we think would happen to the immune response when metabolism was in perfect control (the perfect world scenario) and when it became inconsistent and changeable (the critical illness scenario). In the perfect world scenario, the presence of an insult (e.g., infection or a trauma-related stress signal), would trigger metabolic reprogramming, switching from oxidative phosphorylation to glycolysis. This would enable activation of immune cells and production of pro-inflammatory cytokines and other mediators. At the same time, mitochondrial fission would increase to keep up with the metabolic reprogramming. The slightly elevated mitochondrial ROS and succinate in response to initial insult or cytokines would promote the pro-inflammatory response. Once the insult was eliminated, mitochondrial fusion would increase to create fused elongated mitochondria that favor oxidative phosphorylation and fatty acid oxidation. This would allow activation of regulatory immune cells and production of anti-inflammatory cytokines and other mediators. And itaconate would counteract the effect of succinate, activate the Nrf2-mediated antioxidant pathway to dampen down mitochondrial ROS, and activate ATF3 to inhibit the inflammatory response in macrophages. Immune homeostasis would be achieved as a result. Fig. 1 figure 1
Immunometabolism in the ‘perfect world scenario’ vs. the ‘critical illness scenario’. OXPHOS oxidative phosphorylation, FAO fatty acid oxidation Full size image
In the critical illness scenario, initial metabolic reprogramming from oxidative phosphorylation to glycolysis would go on for longer than necessary, generating excessive lactate (hyperlactatemia) and pro-inflammatory cytokines and mediators. A disrupted mitochondrial fusion/fission cycle could be to blame, one which could not support the timely switch to oxidative phosphorylation and fatty acid oxidation. The anti-inflammatory response would eventually kick in but by then damage would already have occurred to mitochondria and mtDNA because of excessive production of ROS in response to stress or cytokines. Excessive ROS and released mtDNA would aggravate the pro-inflammatory response, which in turn would trigger a more aggressive anti-inflammatory response to try and salvage the situation. The competition between pro- and anti-inflammatory responses would exhaust the nutrients and lead to shutdown of the whole metabolic system. Cells would either die or go into hibernation to preserve energy [26]. This scenario is an over-simplified version of what might happen in the actual disease setting, without considering the crosstalk between cells and organs and many other factors that are not included here. It is designed to shed light on the interaction between the immune response and metabolism. Potential of Mitochondria-Targeting Therapy in Critical Care
Our understanding thus far leads us to think that targeting mitochondria could perhaps correct the underlying cause of immune dysfunction in critical illness and lead to better recovery of the patients. The central role of mitochondrial dynamics in supporting and initiating metabolic reprogramming would make it the perfect therapeutic target. To get the fusion/fission cycle going, the mitochondrial network needs to be replenished by newly synthesized mitochondria via biogenesis. Therapies that could potentially boost mitochondrial biogenesis are mitochondrial transplantation, metformin, nitric oxide (NO), and carbon monoxide. Mitochondrial transplantation has been used successfully in pediatric patients with myocardial ischemia–reperfusion injury [27]. Metformin can activate peroxisome proliferator-activated receptor (PPAR)-gamma coactivator-1α (PGC-1α), and Nrf2, the master regulator of mitochondrial biogenesis and antioxidant systems [28]. Premorbid use of metfor-min is associated with lower mortality in sepsis [29]. NO and carbon monoxide can also enhance mitochondrial biogenesis [30,31,32]. Dietary nitrite has been trialed in patients with coronary artery disease (ClinicalTrials.gov Identifier: NCT00069654). Other therapies, such as mitochondria-targeted antioxidant (MitoQ) [33], could also be beneficial in protecting mtDNA and oxidative phosphorylation from oxidative damage. MitoQ has been trialed in people with Parkinson’s disease (ClinicalTrials. gov Identifier: NCT00329056). Challenges of Applying Mitochondria-Targeting Therapy in Critical Care
There are challenges to overcome before mitochondria-targeting therapy would be possible. First, how do we assess mitochondrial dysfunction in the clinic and identify patients who would benefit from such therapy? A few possible ways could be considered. Non-invasive assessment of mitochondrial oxygen metabolism using a novel device called the COMET monitor was tested on 40 patients during the acute phase of sepsis. This device is based on the protoporphyrin IX-triplet state lifetime technique (PpIX-TSLT) and has been shown to be feasible [33]. This technology is still in its early phase of clinical application but does offer some hope. Another possible biomarker that could potentially be used for assessing mitochondrial dysfunction is plasma mtDNA, but its sensitivity and specificity need further investigation [19, 34, 35]. Furthermore, we could consider using immune response markers as a surrogate markers, one such example could be IFNα inducible protein 27 (IFI27) [36]. If we could overcome the first challenge, the second would be how to deliver mitochondria-targeting therapies to the right organ at the right time. Conclusion
In this chapter, we have demonstrated the important role of mitochondria in regulating the immune response and proposed a scenario that explains immune–metabolism crosstalk in the context of critical illness. We have highlighted the role of mitochon-drial dynamics in overseeing and supporting metabolic reprogramming during immune cell activation. Mitochondrial ROS can be friend or foe when it comes to immune regulation. Two TCA intermediates—succinate and itaconate—with opposite effects have emerged as important players of the immune-regulating mitochon-drial machinery. Our understanding in immunometabolism could take us to the next era of critical care: mitochondria-targeting therapy. Availability of data and material
Not applicable. References
Duggal NA, Snelson C, Shaheen U, Pearce V, Lord JM. Innate and adaptive immune dysregulation in critically ill ICU patients. Sci Rep. 2018;8:10186.
Article
Google Scholar
Surbatovic M, Vojvodic D, Khan W. Immune response in critically ill patients. Mediat Inflamm. 2018;2018:9524315.
Article
Google Scholar
Frazier WJ, Hall MW. Immunoparalysis and adverse outcomes from critical illness. Pediatr Clin N Am. 2008;55:647–68.
Article
Google Scholar
Koutroulis I, Batabyal R, McNamara B, Ledda M, Hoptay C, Freishtat RJ. Sepsis immuno-metabolism: from defining sepsis to understanding how energy production affects immune response. Crit Care Explor. 2019;1:e0061.
Article
Google Scholar
Faas MM, de Vos P. Mitochondrial function in immune cells in health and disease. Biochim Biophys Acta Mol Basis Dis. 2020;1866:165845.
Article
CAS
Google Scholar
Lercher A, Baazim H, Bergthaler A. Systemic immunometabolism: challenges and opportunities. Immunity. 2020;53:496–509.
Article
CAS
Google Scholar
McBride MA, Owen AM, Stothers CL, et al. The metabolic basis of immune dysfunction following sepsis and trauma. Front Immunol. 2020;11:1043.
Article
CAS
Google Scholar
Angajala A, Lim S, Phillips JB, et al. Diverse roles of mitochondria in immune responses: novel insights into immuno-metabolism. Front Immunol. 2018;9:1605.
Article
Google Scholar
Cheng SC, Scicluna BP, Arts RJ, et al. Broad defects in the energy metabolism of leukocytes underlie immunoparalysis in sepsis. Nat Immunol. 2016;17:406–13.
Article
CAS
Google Scholar
Carré JE, Orban JC, Re L, et al. Survival in critical illness is associated with early activation of mitochondrial biogenesis. Am J Respir Crit Care Med. 2010;182:745–51.
Article
Google Scholar
Pearce EL, Pearce EJ. Metabolic pathways in immune cell activation and quiescence. Immunity. 2013;38:633–43.
Article
CAS
Google Scholar
Sack MN. Mitochondrial fidelity and metabolic agility control immune cell fate and function. J Clin Invest. 2018;128:3651–61.
Article
Google Scholar
Haji-Michael PG, Ladrière L, Sener A, Vincent JL, Malaisse WJ. Leukocyte glycolysis and lactate output in animal sepsis and ex vivo human blood. Metabolism. 1999;48:779–85.
Article
CAS
Google Scholar
Gibot S. On the origins of lactate during sepsis. Crit Care. 2012;16:151.
Article
Google Scholar
Zhou R, Yazdi AS, Menu P, Tschopp J. A role for mitochondria in NLRP3 inflammasome activation. Nature. 2011;469:221–5.
Article
CAS
Google Scholar
Agod Z, Fekete T, Budai MM, et al. Regulation of type I interferon responses by mitochondria-derived reactive oxygen species in plasmacytoid dendritic cells. Redox Biol. 2017;13:633–45.
Article
CAS
Google Scholar
Abilés J, de la Cruz AP, Castaño J, et al. Oxidative stress is increased in critically ill patients according to antioxidant vitamins intake, independent of severity: a cohort study. Crit Care. 2006;10:R146.
Article
Google Scholar
Riley JS, Tait SW. Mitochondrial DNA in inflammation and immunity. EMBO Rep. 2020;21:e49799.
Article
CAS
Google Scholar
Harrington JS, Huh JW, Schenck EJ, Nakahira K, Siempos II, Choi AMK. Circulating mitochondrial DNA as predictor of mortality in critically ill patients: a systematic review of clinical studies. Chest. 2019;156:1120–36.
Article
Google Scholar
Murphy MP, O’Neill LAJ. Krebs cycle reimagined: the emerging roles of succinate and itaconate as signal transducers. Cell. 2018;174:780–4.
Article
CAS
Google Scholar
D’Alessandro A, Moore HB, Moore EE, Reisz JA, Wither MJ, Ghasasbyan A, et al. Plasma succinate is a predictor of mortality in critically injured patients. J Trauma Acute Care Surg. 2017;83:491–5.
Article
Google Scholar
Mills EL, Ryan DG, Prag HA, et al. Itaconate is an anti-inflammatory metabolite that activates Nrf2 via alkylation of KEAP1. Nature. 2018;556:113–7.
Article
CAS
Google Scholar
Garbern JC, Lee RT. Mitochondria and metabolic transitions in cardiomyocytes: lessons from development for stem cell-derived cardiomyocytes. Stem Cell Res Ther. 2021;12:177.
Article
Google Scholar
Carelli V, Maresca A, Caporali L, Trifunov S, Zanna C, Rugolo M. Mitochondria: biogenesis and mitophagy balance in segregation and clonal expansion of mitochondrial DNA mutations. Int J Biochem Cell Biol. 2015;63:21–4.
Article
CAS
Google Scholar
Mills EL, Kelly B, O’Neill LAJ. Mitochondria are the powerhouses of immunity. Nat Immunol. 2017;18:488–98.
Article
CAS
Google Scholar
Singer M. The role of mitochondrial dysfunction in sepsis-induced multi-organ failure. Virulence. 2014;5:66–72.
Article
Google Scholar
McCully JD, Cowan DB, Emani SM, Del Nido PJ. Mitochondrial transplantation: from animal models to clinical use in humans. Mitochondrion. 2017;34:127–34.
Article
CAS
Google Scholar
Katila N, Bhurtel S, Park PH, Choi DY. Metformin attenuates rotenone-induced oxidative stress and mitochondrial damage via the AKT/Nrf2 pathway. Neurochem Int. 2021;148:105120.
Article
CAS
Google Scholar
Tan K, Simpson A, Huang S, Tang B, McLean A, Nalos M. The association of premorbid metformin exposure with mortality and organ dysfunction in sepsis: a systematic review and meta-analysis. Crit Care Explor. 2019;1:e0009.
Article
Google Scholar
Nisoli E, Clementi E, Paolucci C, et al. Mitochondrial biogenesis in mammals: the role of endogenous nitric oxide. Science. 2003;299:896–9.
Article
CAS
Google Scholar
Shiva S, Sack MN, Greer JJ, et al. Nitrite augments tolerance to ischemia/reperfusion injury via the modulation of mitochondrial electron transfer. J Exp Med. 2007;204:2089–102.
Article
CAS
Google Scholar
Lancel S, Hassoun SM, Favory R, Decoster B, Motterlini R, Neviere R. Carbon monoxide rescues mice from lethal sepsis by supporting mitochondrial energetic metabolism and activating mitochondrial biogenesis. J Pharmacol Exp Ther. 2009;329:641–8.
Article
CAS
Google Scholar
Lowes DA, Thottakam BM, Webster NR, Murphy MP, Galley HF. The mitochondria-targeted antioxidant MitoQ protects against organ damage in a lipopolysaccharide-peptidoglycan model of sepsis. Free Radic Biol Med. 2008;45:1559–65.
Article
CAS
Google Scholar
Faust HE, Reilly JP, Anderson BJ, et al. Plasma mitochondrial DNA levels are associated with ARDS in trauma and sepsis patients. Chest. 2020;157:67–76.
Article
CAS
Google Scholar
Mao JY, Li DK, Zhang HM, Wang XT, Liu DW. Plasma mitochondrial DNA levels are associated with acute lung injury and mortality in septic patients. BMC Pulm Med. 2021;21:66.
Article
CAS
Google Scholar
Tang BM, Shojaei M, Parnell GP, et al. A novel immune biomarker IFI27 discriminates between influenza and bacteria in patients with suspected respiratory infection. Eur Respir J. 2017;49:1602098.