Hydroxychloroquine (HCQ) Flips from Not Good (June 2021) to Good it Works with Azithromycin (April 2023). 2023-04-09.
During early Pandemic nature scientific reports articles Hydroxychloroquine plus standard of care compared with standard of care alone in COVID-19: a meta-analysis of randomized controlled trials Open Access Published: 07 June 2021 Hydroxychloroquine plus standard of care compared with standard of care alone in COVID-19: a meta-analysis of randomized controlled trials Bahman Amani, Ahmad Khanijahani & Behnam Amani Scientific Reports volume 11, Article number: 11974 (2021) Cite this article 4674 Accesses 12 Citations 41 Altmetric Metrics details Abstract The efficacy and safety of Hydroxychloroquine (HCQ) in treating coronavirus disease (COVID-19) is disputed. This systematic review and meta-analysis aimed to examine the efficacy and safety of HCQ in addition to standard of care (SOC) in COVID-19. PubMed, the Cochrane Library, Embase, Web of sciences, and medRxiv were searched up to March 15, 2021. Clinical studies registry databases were also searched for identifying potential clinical trials. The references list of the key studies was reviewed to identify additional relevant resources. The quality of the included studies was evaluated using the Cochrane Collaboration tool and Jadad checklist. Meta-analysis was performed using RevMan software (version 5.3). Eleven randomized controlled trials with a total number of 8161 patients were identified as eligible for meta-analysis. No significant differences were observed between the two treatment groups in terms of negative rate of polymerase chain reaction (PCR) (Risk ratio [RR]: 0.99, 95% confidence interval (CI) 0.90, 1.08; P = 0.76), PCR negative conversion time (Mean difference [MD]: − 1.06, 95% CI − 3.10, 0.97; P = 0.30), all-cause mortality (RR: 1.09, 95% CI 1.00, 1.20; P = 0.06), body temperature recovery time (MD: − 0.64, 95% CI − 1.37, 0.10; P = 0.09), length of hospital stay (MD: − 0.17, 95% CI − 0.80, 0.46; P = 0.59), use of mechanical ventilation (RR: 1.12, 95% CI 0.95, 1.32; P = 0.19), and disease progression (RR = 0.82, 95% CI 0.37, 1.85; P = 0.64). However, there was a significant difference between two groups regarding adverse events (RR: 1.81, 95% CI 1.36, 2.42; P < 0.05). The findings suggest that the addition of HCQ to SOC has no benefit in the treatment of hospitalized patients with COVID-19. Additionally, it is associated with more adverse events. Today 2023-04-09
Summary Conclusion
Hydroxychloroquine (HCQ) Good or Bad (June 2021). The Jury May Have Just Changed its Mind (April 2023).
WASHINGTON (Reuters) – On January 21, the day the first U.S. case of coronavirus was reported, the secretary of the Department of Health and Human Services appeared on Fox News to report the latest on the disease as it ravaged China. Alex Azar, a 52-year-old lawyer and former drug industry executive, assured Americans the U.S. government was prepared.Centers for Disease Control (CDC) Director Dr. Robert Redfield, U.S. Department of Health and Human Services Chief of Staff Brian Harrison and HHS Secretary Alex Azar meet about the novel coronavirus outbreak in this HHS handout photo taken in Washington, U.S. January 22, 2020. Picture taken January 22. 2020. U.S. Department of Health and Human Services/Handout via REUTERS
“We developed a diagnostic test at the CDC, so we can confirm if somebody has this,” Azar said. “We will be spreading that diagnostic around the country so that we are able to do rapid testing on site.”
While coronavirus in Wuhan, China, was “potentially serious,” Azar assured viewers in America, it “was one for which we have a playbook.”
Azar’s initial comments misfired on two fronts. Like many U.S. officials, from President Donald Trump on down, he underestimated the pandemic’s severity. He also overestimated his agency’s preparedness.
As is now widely known, two agencies Azar oversaw as HHS secretary, the Centers for Disease Control and Prevention and the Food and Drug Administration, wouldn’t come up with viable tests for five and half weeks, even as other countries and the World Health Organization had already prepared their own.
Shortly after his televised comments, Azar tapped a trusted aide with minimal public health experience to lead the agency’s day-to-day response to COVID-19. The aide, Brian Harrison, had joined the department after running a dog-breeding business for six years. Five sources say some officials in the White House derisively called him “the dog breeder.”
Azar’s optimistic public pronouncement and choice of an inexperienced manager are emblematic of his agency’s oft-troubled response to the crisis. His HHS is a behemoth department, overseeing almost every federal public health agency in the country, with a $1.3 trillion budget that exceeds the gross national product of most countries.
Azar and his top deputies oversaw health agencies that were slow to alert the public to the magnitude of the crisis, to produce a test to tell patients if they were sick, and to provide protective masks to hospitals even as physicians pleaded for them.
The first test created by the CDC, meant to be used by other labs, was plagued by a glitch that rendered it useless and wasn’t fixed for weeks. It wasn’t until March that tests by other labs went into production. The lack of tests “limited hospitals’ ability to monitor the health of patients and staff,” the HHS Inspector General said in a report this month. The equipment shortage “put staff and patients at risk.”
A promised virus surveillance program failed to take root, despite assurances Azar gave to Congress. Rather than share information, three current and three former government officials told Reuters, Azar and top staff sidelined key agencies that could have played a higher-profile role in addressing the pandemic. “It was a mess,” said a White House official who worked with HHS.
Officials across the government, from President Trump on down, have been blasted for America’s halting response to the pandemic. Critics inside and outside the administration say a meaningful share of the responsibility lies with HHS and Trump appointee Azar.
“You have to blame the problem on the virus, but it’s Azar’s operation,” said Lynn Goldman, the dean of the public health school at George Washington University, who has served on advisory boards of the FDA and CDC. “And the buck stops there.”
HHS declined to make Azar available for an interview. Michael Caputo, the new chief HHS spokesman, declined to answer Reuters questions about Azar’s stewardship, saying in a statement: “We are communicating to the American public during a deadly pandemic.”
DALLAS LABRADOODLES
Azar is a Republican lawyer who once clerked for the late conservative Supreme Court Justice Antonin Scalia and counts current Supreme Court Justice Brett Kavanaugh as a friend. Under George W. Bush, Azar worked for HHS as general counsel and deputy secretary. During the Obama years, he cycled through the private sector as a pharmaceutical company lobbyist and executive for Eli Lilly. After Trump’s first HHS secretary was forced out in a travel corruption scandal, Azar stepped in, in January 2018.
Two years later, at the dawn of the coronavirus crisis, Azar appointed his most trusted aide and chief of staff, Harrison, as HHS’s main coordinator for the government’s response to the virus.
Harrison, 37, was an unusual choice, with no formal education in public health, management, or medicine and with only limited experience in the fields. In 2006, he joined HHS in a one-year stint as a “Confidential Assistant” to Azar, who was then deputy secretary. He also had posts working for Vice President Dick Cheney, the Department of Defense and a Washington public relations company.
Before joining the Trump Administration in January 2018, Harrison’s official HHS biography says, he “ran a small business in Texas.” The biography does not disclose the name or nature of that business, but his personal financial disclosure forms show that from 2012 until 2018 he ran a company called Dallas Labradoodles.
The company sells Australian Labradoodles, a breed that is a cross between a Labrador Retriever and a Poodle. He sold it in April 2018, his financial disclosure form said. HHS emailed Reuters that the sale price was $225,000.
At HHS, Harrison was initially deputy chief of staff before being promoted, in the summer of 2019, to replace Azar’s first chief of staff, Peter Urbanowicz, an experienced hospital executive with decades of experience in public health.
This January, Harrison became a key manager of the HHS virus response. “Everyone had to report up through him,” said one HHS official.Slideshow ( 2 images )
One questionable decision, three sources say, came that month, after the White House announced it was convening a coronavirus task force. The HHS role was to muster resources from key public health agencies: the CDC, FDA, National Institutes of Health, Office of Global Affairs and the Assistant Secretary for Preparedness and Response.
Harrison decided, the sources say, to exclude FDA Commissioner Stephen Hahn from the task force. “He said he didn’t need to be included,” said one official with knowledge of the matter.
When task force members were announced January 29, neither Hahn nor the FDA were included. Hahn wasn’t put on the task force until Vice President Mike Pence took over in February. Two of Hahn’s high-profile counterparts were on it from the start: CDC director Robert Redfield and Dr. Anthony Fauci, director of the National Institute of Allergy and Infectious Diseases.
The HHS denied it was Harrison’s decision to leave out Hahn and the FDA, but declined to say who made the call. The agency lauded Harrison’s work on the task force.
In a statement, Hahn said the FDA was focused on the coronavirus epidemic, “not on when we were added to the task force,” and that the agency was not “excluded.”
Fauci, who has become a public face of the Trump Administration’s COVID-19 effort, said he wasn’t sure including the FDA was necessary at the start. Initially, the Chinese government was saying the virus spread through animals, not human to human, he said. “You would include the FDA when you want to expedite drugs or devices,” Fauci said.
Others said the lack of a strong FDA role early on had direct consequences. Two sources familiar with events say the White House wasn’t getting information from the FDA about the state of the testing effort, a crucial element of the coronavirus response.
Reached by phone, Harrison declined to answer Reuters’ questions. In a later statement, he did not address questions about the task force but said he was proud of his work history. “Americans would be well served by having more government officials who have started and worked in small family businesses and fewer trying to use that experience to attack them and distort the record,” he wrote.
In a statement to Reuters, Azar said Harrison has been an asset. “From day one, Brian has demonstrated remarkable leadership and managerial talents,” Azar wrote.
LOW RISK?
In the pandemic’s early days, Azar offered words of both concern and assurance in public. On January 31, a day after the WHO declared COVID-19 a global health emergency, Azar declared it a public health emergency.
That same day, during the first Coronavirus Task Force briefing, Azar told the public: “I want to stress: The risk of infection for Americans remains low.”
The United States, he said, had taken adequate precautions. Travel restrictions and 14-day quarantines on Americans who had been to Wuhan, where the virus originated, were imposed. Americans returning from other parts of China had to self-quarantine.
The next week, on February 7, in another press conference, Azar repeated the message. “The immediate risk to the American public is low at this time,” he announced.
Behind the scenes, his aides say, Azar had alerted the White House in early January, and then later that month spoke directly to the president. It is unclear exactly what Azar told the president, because transcripts are not available.
“There’s a lot of CYA going on,” said one senior administration official, who said Azar never spelled out that stockpiles of protective equipment might be inadequate or the tests were not working. “We were told the test was ready. That turned out to be flat-out wrong.”
Trump denied Azar sent out alarms. “@SecAzar told me nothing until later,” he tweeted earlier this month.
Meanwhile, Azar continued to say “the immediate risk” to Americans was low and that travel restrictions had worked. “So I think so far, our measures have been quite effective,” he told NPR on February 14.
Others were raising alarms. “It’s not so much of a question of if this will happen any more, but rather more of a question of exactly when this will happen,” Dr. Nancy Messonnier, director of the National Center for Immunization and Respiratory Diseases, said at a February 25 news briefing.
MORE GLITCHES
Responding to Congressional concerns, Azar said HHS had launched a coronavirus surveillance system in five cities. The plan was to test patients who showed up with flu symptoms, to see if they actually were infected with the novel coronavirus.
But the system was either delayed or not implemented in the cities and now is seen by epidemiologists as irrelevant given the massive community spread and continued inadequate testing.
By the end of February, Azar sought more money to attack the crisis as he testified before Congress. “This is an unprecedented potentially severe health challenge globally, and will require additional measures,” he said.
Still, he assured senators his agency was in control. “We have enacted the most aggressive containment measures in the history of our country,” he said.
He again provided words of calm, appearing on Fox News. “But thanks to President Trump’s historically aggressive containment efforts, we’ve actually contained the spread of this virus here in the United States at this point,” he said February 25. “I think part of the message to the American people is we all need to take a bit of deep breath here.”
“The government is working on this. You’ve got the right people on this.”
By the end of February, Azar and Harrison were no longer running the White House task force. That month, Vice President Pence took control. The FDA and Hahn are now actively involved. A Pence spokesperson said the issue of precluding the FDA from the task force “pre-dates the VP’s leadership” and declined further comment.
Azar seemed caught off guard by the change. “I’m still chairman of the task force,” he told the press after Pence took over.
Given Azar’s early struggles, the White House should have taken a stronger role over the task force from the outset, said Ashish Jha, director of Harvard University’s Global Health Institute. “It was very clear that Azar wasn’t able to marshal the forces across the government like he needed to,” he said.
Jeffrey Flier, a former Harvard Medical School dean, said the HHS role remains as vital as ever. As of Wednesday, over 47,000 Americans have died of COVID-19, and more than 830,000 have been infected.
“Clearly there was a need for better coordination of the FDA and CDC and other agencies,” he said. “HHS has to be operating effectively in a crisis like this.”
Reporting by Aram Roston and Marisa Taylor in Washington. Editing by Ronnie Greene
HOW THEY DID IT: They used a particular bug (Coronavirus) that was previously relatively benign and nonpathogenic and modified it through the use of gene editing techniques to make the bug (Covid-19) virulent, pathogenic, dangerous and then released that bug (Covid) in key sites.… Show mor
THEY RELEASED COVID-19: U.S. Patents show CDC ownership of Coronavirus. Both China and the U.S. involved in creating SARS-CoV-2. Bill Gates and CCP appoints criminal Tedros of WHO. The DOD, FDA, CIA, NIH, Fauci, Baric, Daszak, Rockefeller, Rothschilds are all involved in these Crimes.
Bill Gates and the Rockefeller foundation paid Google, Facebook, Politico, Wikipedia, Fact Checkers in order to censor and control all the information. The CIA has been using Operation Mockingbird for years and has over 3,000 agents implanted in Mainstream Media to control the population. Event 201 was sponsored by Bill Gates, the Johns Hopkins Center for Health Security (CIA) and the World Economic Forum to enforce a worldwide Pandemic response 5 months before the WHO fraudulently declared a global pandemic.
It was a planned coordinated criminal effort worldwide. In January 2017 Anthony Fauci said there will be a surprise virus outbreak before the end of 2020. Bill Gates in 2015 talked of a future pandemic and lied in April 2020 when he said they did not simulate or practice for a pandemic.
Klaus Schwab in his book Covid-19 The Great Reset shows Covid was the Trojan Horse to Reset the World according to the UN 2030 Agenda. Build Back Better slogan is a criminal coordinated effort to remove human rights and institute a one world government.
Bill Gates and the Rockefeller foundation bribes the WHO, NIH, NIAID, CDC, FDA, Medical Schools and Journals to control the health industry and public health policy.
WHO Chief Tedros involved in genocide killing and torture in Ethiopia. Tedros is a known member of the communist party. He is Beijing’s and Gates puppet. As a Health Minister he was accused of covering up three Cholera Epidemics and committing crimes against humanity. CCP and Bill Gates helped put Tedros in charge of the WHO. John D. Rockefeller over 100 years ago seized the U.S. Media and took control over public health using toxic petroleum based drugs for profit and controlled the American Medical Association blacklisting and expelling any doctors who practiced natural medicine. Rockefeller’s poison injections and medicines started causing cancer in early years and to cover it up formed the American Cancer Society. Medical error is the 3rd leading cause of death in America.
Bill Gates used India and Africa as guinea pigs for pharmaceutical companies to make a financial killing while killing a lot of people in the process including killing innocent children and babies with vaccines. Bill Gates controls GAVI The Vaccine Alliance to vaccinate the world with his poisons.
National Security Study Memorandum NSSM 200 Implications of Worldwide Population Growth For U.S. Security and Overseas Interests December 10, 1974 (THE KISSINGER REPORT) shows the intention of governments to reduce the population. Bill Gates is one of the key funders in the Stratosphere experiment to block out the sun for Climate Change by releasing poisons in the air. Environmental Scientist call it global genocide experiment.
Gates has invested over one billion dollars in the Earth Now Global Surveillance project to launch hundreds of satellites to monitor people everywhere 24/7 a day. In partnership with MIT Bill Gates has developed a new technology that allows vaccines to be injected under your skin along with your medical records. Bill Gates Gates funded genetically modified mosquitoes released in the USA to allow human immunization by means of mosquito bites “Flying Syringes.” Gates had business dealings and a relationship with Jeffrey Epstein, a convicted child sex criminal. Why would he choose to partner with the world’s most notorious pedophile? Gates is the top financial donor of the WHO and CDC. No one person has more power than Gates to influence and control the health and medical freedom of all people. Bill Gates and all mRNA Vaccines must be stopped. This is a global genocide experiment and a takeover of humanity.
THEY RELEASED COVID-19: U.S. Patents show CDC ownership of Coronavirus. Both China and the U.S. involved in creating SARS-CoV-2. Bill Gates and CCP appoints criminal Tedros of WHO. The DOD, FDA, CIA, NIH, Fauci, Baric, Daszak, Rockefeller, Rothschilds are pic.twitter.com/POFAcPf7Yo…
Reference: Ghielmetti M, Schaufelberger HD, Mieli-Vergani G, Cerny A, Dayer E, Vergani D, Terziroli Beretta-Piccoli B. Acute autoimmune-like hepatitis with atypical anti-mitochondrial antibody after mRNA COVID-19 vaccination: A novel clinical entity? J Autoimmun. 2021 Sep;123:102706. doi: 10.1016/j.jaut.2021.102706. Epub 2021 Jul 15. PMID: 34293683; PMCID: PMC8279947.
Abstract
Autoimmune phenomena and clinically apparent autoimmune diseases, including autoimmune hepatitis, are increasingly been reported not only after natural infection with the SARS-CoV-2 virus, but also after vaccination against it. We report the case of a 63-year old man without a history of autoimmunity or SARS-CoV-2 natural infection who experienced acute severe autoimmune-like hepatitis seven days after the first dose of the mRNA-1273 SARS-CoV-2 vaccine. Liver histology showed inflammatory portal infiltrate with interface hepatitis, lobular and centrilobular inflammation with centrilobular necrosis, in absence of fibrosis and steatosis. Serum immunoglobulin G was slightly elevated. Autoimmune liver serology showed an indirect immuno fluorescence pattern on triple rodent tissue compatible with anti-mitochondrial antibody (AMA), but, unexpectedly, this pattern was not mirrored by positivity for primary biliary cholangitis (PBC)-specific molecular tests, indicating that this antibody is different from classical AMA. Anti-nuclear antibody (ANA) was also positive with a rim-like indirect immuno fluorescence pattern on liver and HEp2 cell substrates, similar to PBC-specific ANA; however, anti-gp210 and a large panel of molecular-based assays for nuclear antigens were negative, suggesting a unique ANA in our patient. He carries the HLA DRB1*11:01 allele, which is protective against PBC. Response to prednisone treatment was satisfactory. The clinical significance of these novel specificities needs to be further evaluated in this emerging condition.
Jyrkkanen Comment: The mRNA-1273 SARS-CoV-2 vaccine vaccination appears to have induced anti-mitochondrial antibody AMA and anti-nuclear antibody ANA, suggesting the primary support genetic reservoirs of the host had turned antigenic and become autoimmune targets for attack by the hosts immune system. I suspect it is from spike protein. This autoimmune reaction would lead to the destruction of the source of ATP+NAD production and immune system integrity and would lead to and increased risk of cancer, heart disease and other medical conditions.
CONSEQUENCES OF LOSS OF A VACCINATED POPULATIONS’ IMMUNE INTEGRITY MIGHT BE BE AN INCREASE IN RARE DISEASES EX: MONKEY POX IN ISRAEL
Twitter Censors Pfizer-Injured Israeli COVID Vaccine Director
Prof. Shmuel Shapira MD MPH (Col.), who served as Director of the Israel Institute for Biological Research between 2013 and 2021, suggested that the monkeypox outbreak was connected to mRNA vaccines.
Aug 3, 2022
Professor Shmuel Shapira, M.D., MPH, served as the Director General of the Israel Institute for Biological Research (IIBR) between 2013 and 2021, where he led Israel’s effort to develop a coronavirus vaccine.
Prof. Shapira is also the founder and head of the Department of Military Medicine of the Hebrew University Faculty of Medicine and IDF Medical Corps. He is a Senior Research Fellow at the International Institute for Counter-Terrorism (ICT) at Reichman University in Israel.
Shapira previously served as Deputy Director General of the Hadassah Medical Organization and as the Director of the Hebrew University Hadassah School of Public Health. He is a Full Colonel (Res.) in the Israel Defense Forces (IDF) and served as the IDF Head of Trauma Branch.
He has published more than 110 peer-reviewed scientific articles and is the editor of Essentials of Terror Medicine, Best Practice for Medical Management of Terror Incidents, and Medical Response to Terror Threats.
Last week, Twitter censored Prof. Shapira—who was “physically injured” after his third Pfizer vaccine—and forced him to remove a post which said: “Monkey pox cases were rare for years. During the last years a single case was documented in Israel. It is well established the mRNA vaccines affect the natural immune system. A monkey pox outbreak following massive covid vaccination: *Is not a coincidence.”
Antibiotics Use in Hospitalised COVID-19 Patients in a Tertiary Care Centre: A Descriptive Cross-sectional Study and Heart Mitochondria. JORMA JYRKKANEN 2023-03-23
Thapa B, Pathak SB, Jha N, Sijapati MJ, Shankar PR. Antibiotics Use in Hospitalised COVID-19 Patients in a Tertiary Care Centre: A Descriptive Cross-sectional Study. JNMA J Nepal Med Assoc. 2022 Jul 1;60(251):625-630. doi: 10.31729/jnma.7394. PMID: 36705203; PMCID: PMC9297358.
ABSTRACT Introduction:
Antimicrobial resistance is a global health problem. The widespread and improper antibiotics use is the leading cause of antimicrobial resistance. Bacterial co-infection in COVID-19 patients is the basis for the use of antibiotics in the management of COVID-19. COVID-19 pandemic has seriously impacted antibiotic stewardship and increased the global usage of antibiotics, worsening the antimicrobial resistance problem. The use of antibiotics among COVID-19 patients is high but there are limited studies in the context of Nepal. This study aimed to find out the prevalence of antibiotic use among hospitalised COVID-19 patients in a tertiary care centre.
Introduction:
Antimicrobial resistance is a global health problem. The widespread and improper antibiotics use is the leading cause of antimicrobial resistance. Bacterial co-infection in COVID-19 patients is the basis for the use of antibiotics in the management of COVID-19. COVID-19 pandemic has seriously impacted antibiotic stewardship and increased the global usage of antibiotics, worsening the antimicrobial resistance problem. The use of antibiotics among COVID-19 patients is high but there are limited studies in the context of Nepal. This study aimed to find out the prevalence of antibiotic use among hospitalised COVID-19 patients in a tertiary care centre. Methods:
A descriptive cross-sectional study was conducted on hospitalised COVID-19 patients from April 2021 to June 2021 in a tertiary care centre. Ethical approval was taken from the Institutional Review Committee (Reference number: 2078/79/05). The hospital data were collected in the proforma by reviewing the patient’s medical records during the study period of 2 months. Convenience sampling was used. Point estimate and 95% Confidence Interval were calculated. Results:
Among 106 hospitalised COVID-19 patients, the prevalence of antibiotics use was 104 (98.11%) (95.52-100, 95% Confidence Interval). About 74 (71.15%) of patients received multiple antibiotics. The most common classes of antibiotics used were cephalosporins, seen in 85 (81.73%) and macrolides, seen in 57 (54.81%) patients. Conclusions:
The prevalence of antibiotics use among hospitalised COVID-19 patients was found to be higher when compared to other studies conducted in similar settings. Keywords: antibiotics, bacterial infection, co-infection, COVID-19 Go to: INTRODUCTION
Antimicrobial resistance (AMR) is a major threat to global public health due to the increasing incidence of resistant human pathogens.1,2 The widespread and improper use of antibiotics is the leading cause of AMR.1 Coronavirus Disease 2019 (COVID-19) is a viral disease thus untreatable by antibiotics, but the viral respiratory infections may clinically progress to bacterial pneumonia requiring antibiotic administration.2 This co-pathogenesis is the basis for use of antibiotics in COVID-19. But appropriate use of antibiotics is utmost to prevent AMR.
COVID-19 pandemic has seriously impacted antibiotic stewardship and single-handedly increased the global usage of antibiotics, causing a cascading effect on the AMR problem. The use of antibiotics among COVID-19 patients is high but there are limited studies in the context of Nepal.3-5
This study aimed to find out the prevalence of antibiotics use among COVID-19 patients of a tertiary care centre. Go to: METHODS
A descriptive cross-sectional study was conducted at KIST Medical College and Teaching Hospital after taking ethical approval from the Institutional Review Committee (Reference number: 2078/79/05). The study was conducted during the study period from 6 August 2021 to 6 October 2021 during which hospitalised COVID-19 patients admitted from April 2021 to June 2021 were studied. All the COVID-19 cases confirmed by reverse transcriptase polymerase chain reaction (RT-PCR) test who were admitted in the dedicated COVID-19 ward, high dependency unit (HDU) and intensive care units (ICU) were enrolled. Patients who had incomplete documentation were excluded from the study. Convenience sampling was used. The sample size was calculated using the following formula: n=Z2×p×qe2=1.962×0.50×0.500.102=97
Where,
n = minimum required sample size
Z = 1.96 at 95% Confidence Interval (CI)
p = prevalence taken as 50% for maximum sample size calculation
q = 1-p
e = margin of error, 10%
Minimum sample size calculated was 97. However, we enrolled 106 cases. The collected data from hospital records was entered in the proforma by reviewing the patient’s medical records during the study period of two months. Demographic profile of patients like age and sex, clinical profile like co-morbidity and disease severity, management profile like level of care required for patients’ treatment, number and type of antibiotics used, route of antibiotic administration, duration of antibiotics used, and estimated cost of antibiotics used for the treatment were assessed. The patients who were treated with at least one antibiotic were included. All the cases were classified as a mild disease, moderate disease, or severe disease.6
In our study, 17 different antibiotics were used belonging to seven different antibiotic classes. They are namely cephalosporin (ceftriaxone, cefixime, cefoperazone, and cefepime), macrolides (azithromycin, clindamycin, and erythromycin), penicillin group (piperacillin, amoxicillin), quinolones (moxifloxacin, levofloxacin, ciprofloxacin), imidazoles (metronidazole), carbapenem (meropenem) and beta-lactamase inhibitors (clavulanic acid, tazobactam, sulbactam).
The data were entered and analysed using IBM SPSS Statistics 21.0. Point estimate and 95% CI were calculated. Go to: RESULTS
Among 106 hospitalised COVID-19 patients, the prevalence of use of antibiotics was 104 (98.11%) (95.52-100, 95% CI). The mean number of antibiotics used per patient was 1.86 ±0.64. A total of 74 (71.15%) patients were under two or more antibiotic therapy. Around 60 (57.69%) patients were treated with intravenous as well as per oral route of administration of antibiotics.
The mean number of days of admission was 6.44±4.81 days. The mean duration of antibiotics use was 6.33 ±2.72 days. About 30 (28.85%) received a 5 day course of antibiotics while 25 (24.04%) patients received a 7 days course of antibiotics. Only 23 (22.12%) received antibiotic therapy for more than 7 days. The mean estimated expenditure on antibiotics was NPR 4,645±8, 498 (USD 38.71±70.82) (Table 1). Table 1 Use of antibiotics in management of COVID-19 patients (n = 104). Characteristics n (%) Number of antibiotics used 1 30 (28.85) 2 59 (56.73) 3 15 (14.42) Route of antibiotics Intravenous route only 37 (35.58) Per oral route only 7 (6.73) Intravenous and per oral route 60 (57.69) Total number of days of antibiotics use ≤7 days 81 (77.88)
7 days 23 (22.12) Estimated expenditure on antibiotics therapy NPR (US dollar) ≤1,200 (USD 10) 43 (41.35) 1201 to 6,000 (USD 11 to 50) 42 (40.38) 6,001 to 12, 000 (USD 51 to 100) 8 (7.69) 12,001 to 24,000 (USD 101 to 200) 7 (6.73) 24,000 (USD 200) 4 (3.85) Open in a separate window
The mean age of the patients was 55.84±18 years. A total of 54 (51.92%) patients were males. Around 59 (56.73%) had at least one comorbid condition with the most common conditions being hypertension seen in 39 (37.50%) and diabetes mellitus seen in 23 (22.16%) (Table 2). Table 2 Demographic characteristics of hospitalised COVID-19 patients who received antibiotic therapy (n= 104). Age group n (%) ≤20 years 5 (4.81) 21 to 40 years 18 (17.31) 41 to 60 years 37 (35.58) 61 to 80 years 37 (35.58)
80 years 7 (6.73) Sex Males 54 (51.92) Females 50 (48.08) Comorbidities Diabetes mellitus 23 (22.16) Hypertension 39 (37.50) Chronic Obstructive Pulmonary 10 (9.62) Disease (COPD) Hypothyroidism 12 (11.54) Psychiatric illness 2 (1.92) Heart failure 2 (1.92) Autoimmune disease 2 (1.92) Chronic kidney disease 1 (0.96) Open in a separate window
Severe COVID-19 represented 37 (35.58%) of total patients, 16 (15.38%) of them were managed in ICU with ventilator support. Moderate COVID-19 cases also accounted for 37 (35.58%) of total patients. These patients were mostly managed in a dedicated COVID-19 ward with 2 (1.92%) cases managed in ICU and 2 (1.92%) in HDU. All 30 (28.84%) mild cases were managed in the ward (Table 3). Table 3 COVID-19 severity and level of care received by the patients who received antibiotic therapy (n = 104). Level of care Mild n (%) Moderate n (%) Severe n (%) Total n (%) ICU with ventilator support – – 16 (15.38) 16 (15.38) ICU without ventilator support – 2 (1.92) 4 (3.85) 6 (5.77) HDU – 2 (1.92) 15 (14.42) 17 (16.35) Ward 30 (28.84) 33 (31.73) 2 (1.92) 65 (62.50) Open in a separate window
The most common class of antibiotics used was cephalosporins in 85 (81.73%) patients followed by macrolides in 57 (54.81 %). Cefixime used in all cases was a substitute for ceftriaxone in oral form in 18 (17.31%) patients who were previously prescribed ceftriaxone. Beta-lactamase inhibitors were used in 35 (33.65%) in conjunction with penicillin (amoxicillin) or cephalosporin group of drugs (cefoperazone, cefepime). The most common combination used was cephalosporin with macrolides at 38 (36.54%) (Table 4). Table 4 Types of antibiotics used in management of COVID-19 patients (n = 104). Antibiotics n (%) Cephalosporins prescribed parenterally 85 (81.73) Ceftriaxone 76 (73.08) Cefepime sulbactam 3 (2.88) Cefoperazone sulbactam 6 (5.77) Cephalosporins prescribed enterally 18 (17.31) Cefixime 18 (17.31) Macrolides 57 (54.81) Azithromycin 55 (52.88) Erythromycin 1 (0.96) Clindamycin 1 (0.96) Penicillins 26 (25.00) Piperacillin tazobactam 14 (13.46) Amoxicillin clavulanic acid 12 (11.54) Quinolones 11 (10.58) Moxifloxacin 9 (8.65) Levofloxacin 2 (1.92) Ciprofloxacin 1 (0.96) Imidazoles 10 (9.62) Metronidazole 10 (9.62) Carbapenems 4 (3.85) Meropenem 4 (3.85) THIS IS A COCKTAIL THAT IS VERY DANGEROUS TO HEART MITOCHONDRIA-JORMA JYRKKANEN COMMENT DISCUSSION
The prevalence of use of antibiotics was 98.1%. About 71.15% patients were treated with two or more antibiotics. The mean number of antibiotics used per patient was 1.86. The mean duration of antibiotics use was 6.33 days. Seventeen different antibiotics were used belonging to 7 different antibiotic classes. The most common class of antibiotics used was cephalosporin at 85 (81.73%) and macrolides at 57 (54.81%).
Even before the COVID-19 pandemic, AMR was projected to become responsible for approximately 10 million deaths worldwide in the coming three decades.7 COVID-19 has undoubtedly affected antibiotic stewardship and has increased antibiotic consumption patterns globally, adding to the already existing global AMR problem. Because of this, the mortality due to AMR is expected to be higher in post COVID era.2 This pandemic has disrupted health delivery systems worldwide. This has increased the overuse of antibiotics, eventually leading to resistant organisms requiring aggressive treatment.8 Thus AMR is a problem of greater concern than COVID-19 which has unfortunately been overshadowed amidst the pandemic.7,9 Increased use of antibiotics is more challenging, especially in the low and middle-income countries (LMIC) due to the inefficiency and inadequacy of health care services.2
The Infectious Diseases Society of America (IDSA) states that only 8% of the COVID-19 patients acquired bacterial/fungal superinfections requiring antibiotics.10 However, a study showed 72% of COVID-19 patients received empirical broad-spectrum antibiotics, even when bacterial coinfection was absent.11 Current World Health Organization guidelines indicate that antibiotics should not be prescribed in mild or moderate COVID-19 cases unless there are pre-existing symptoms of bacterial co-infection. Furthermore, when treating severe cases with an empirical antimicrobial agent, the overall condition of the patient, local bacterial epidemiology, and clinical judgement should be integrated, to ensure judicial antimicrobial usage.12 In COVID-19 patients, antibiotics are used for potential anti-inflammatory, immune-modulating, and potential antiviral properties. But the antiviral mechanism of these agents is doubtful. This widespread antibiotic use is likely to worsen preexisting AMR crisis.13
The influenza pandemic was largely a problem of viral infection complicated by bacterial co-pathogenesis.14 This has been our basis for use of a wide range antibiotics empirically though COVID-19 is primarily a viral pathology and is not conventionally treated with antibiotics.
In a study, 71.00% of the hospitalised COVID-19 patients received antibiotics despite a confirmed bacterial co-infection rate of only 1%.3 Antibiotic was used in 95.00% COVID-19 patients when secondary bacterial infection was only found in 15.00%.4 A systematic review showed the mean rate of antibiotic use was 74.00 %.5 In our study, the prevalence of use of antibiotics was 98.10% which is very high when compared to above studies. In most cases antibiotics use often empirical. Empiric antibiotics were often used for the concern of community-acquired pneumonia (89.00%).15 This showed that antibiotic therapy has been used often empirically in the majority of patients even when very few were proven to have bacterial coinfection.
In our study, 17 different antibiotics belonging to seven antibiotic classes were used. Similar to our study a wide range of antibiotics use was documented in other studies.1,5,10,13,15-18 Many other classes of antibiotics other than above were used in other studies for the management of COVID-19 patients. They are aminoglycosides,1 glycopeptide antibiotic like vancomycin and teicoplanin,10 oxazolidinones like linezolid, tetracycline and cyclic lipopeptides like daptomycin.17 Most of these are newer classes of antibiotics and increased use of these should raise a red flag among concerned clinicians, pharmacists, microbiologists, public health experts, hospitals, local authorities as well as regulatory bodies.
Carbapenem, fluoroquinolones, and aminoglycoside were highly prevalent in ICU patients.1 Similar to this carbapenem was exclusively used for ICU patients in our study. Other commonly used antibiotics among ICU patients were fluoroquinolones, cephalosporin, piperacillin with tazobactam, and macrolides. In general, ICU addmission compromises of a very sick patient with superadded bacterial infection and in regard to COVID-19, it comprises of severe COVID-19 infection often requiring ventilatory support. In such conditions, it is common practice to use multiple higher and broad-spectrum antibiotics.
The common antibiotics in use were ceftriaxone (54.00%), vancomycin (48.00%), azithromycin (47.00%), and cefepime (45.00%).10 In our study ceftriaxone (73.08%) and azithromycin (52.88%) were widely used but cefepime was used in 2.88% of patients and vancomycin was not used at all. Higher antibiotics like cefepime and vancomycin should only be used when there is a valid indication, otherwise, it may result in resistant infection which will be very hard to treat.
Ceftriaxone and azithromycin are often the most common antibiotics used in the management of COVID-19 patients.10,13,15,16 Most of the local guidelines as well as some international guidelines advocate for use of these antibiotics based on the epidemiology of local pathogens and resistance patterns. The advantage of the use of these antibiotics is that it covers most of the opportunistic pathogens that could cause secondary infection in COVID-19. But on the other hand, wide and inappropriate use of these antibiotics can lead to with emergence resistance of these common, cheap, and very efficient antibiotics.
Macrolide, specifically azithromycin, was the most common antibiotic used in the clinical management of COVID-19.13 Macrolides, particularly azithromycin, were used in the treatment of more than half of the patients in our study. These drugs are often used to cover atypical organism that have the potential to cause a secondary infection.10,11
Fluoroquinolones were most used, (56.80%), followed by ceftriaxone (39.50%), then azithromycin (29.10%), and carbapenems were only used in two patients.18 Unlike to above study, in our study fluoroquinolones were used less (10.58%) and ceftriaxone and azithromycin was basically used in most patients. But similarity was observed between ours and the above study regarding the use of carbapenems, which was used in 3.85% of patients. Wide use of carbapenem, used in up to 40.10% of patients was also reported.5 carbapenem is often used as a reserved antibiotic for severe infection thus minimal use of these in COVID-19 signifies the presence of good antibiotics stewardship and antibiotic management system among concerned institutions while wide use can signify the opposite.
The most common antibiotic used was third-generation cephalosporin (ceftriaxone) (53.80%), moxifloxacin (29.50%), and doxycycline (25.40%).5 Similar to the above study the most common class of antibiotics used in our study was cephalosporin (81.74%), around 90.00% of which was third-generation cephalosporin namely ceftriaxone (76/85). But unlike the above, Moxifloxacin was used in only 8.65% and doxycycline or other tetracycline group of drugs was not used at all in our study.
A study showed all patients were receiving at least one antibiotic with 31.08% receiving a single antibiotic and 68.91% receiving multiple antibiotics.5 Similar to this study, 98.10% of patients in our study received at least one antibiotic, 28.35% received single antibiotic agents and 71.15% received multiple antibiotics. But the mean number of antibiotics used in the study above was 2.02 which is more when compared to 1.85 in our study. In 34.6%, three antibiotics were given simultaneously while 9.6% received only one antibiotic.16 Unlike the above, in our study three antibiotics were used in only 14.42% and a single antibiotic was used in 28.85%. The use of multiple antibiotics is worrisome as this might represent unchecked use of antibiotics which will contribute to the worldwide problem of AMR.
In our study, patients with comorbidity were found to have received multiple antibiotics (72.90%). Around 74.00% of patients with diabetes were on multiple antibiotics. Similar findings were documented in another study.5 This might be due to COVID-19 patients with comorbidities like diabetes, airway diseases, hypertension being at greater risk of developing secondary bacterial infection.19 After predicting the risk of secondary infection, multiple antibiotics were used empirically in those patients.
Overall, patients presenting with severe disease received more antibiotics.5 This is also true for our study. COVID-19 patients with co-morbidities and severe COVID-19 are the two most vulnerable groups of patients, thus multiple antibiotics have been found to be used liberally in these patients. These situations can be dealt with systematically by establishing standard antibiotic prescribing guidelines considering local pathogens and sensitivity patterns to antibiotics. This could be further reinforced by appropriate clinical knowledge, laboratory facilities, and surveillance systems.
In our study the mean duration of antibiotics treatment was 6.33 days which is nearly half when compared to 12.71 days with a range from 3 days to 23 days.16 Similar result was found in another study.18 Both of these, decreased and extended duration of antibiotic treatment might represent inappropriate and improper use of antibiotics regimen. Because both, underuse and overuse of antibiotics can result in the emergence of resistance.
Our study is a single centred study and has a small sample size. Therefore, our findings may not be generalizable to other settings. Go to: CONCLUSIONS
The prevalence of use of antibiotics among hospitalised COVID-19 patients was found to be higher when compared to other studies conducted in similar settings. Potential bacterial co-infection has been the basis for the use of antibiotics in the management of COVID-19 patients. The rate and number of antibiotics used for mild to moderate disease were also high. The common class of antibiotics used are cephalosporin and macrolides namely ceftriaxone and azithromycin. Higher class antibiotics were mostly used in the management of severe disease in ICU and with ventilator support. However, judicial use of antibiotics among COVID-19 patients with variable severity especially among those admitted in ICU and on ventilatory support could be promoted in order to reduce AMR during this COVID-19 pandemic. Robust antibiotic stewardship programs and surveillance systems should be implemented. Go to: ACKNOWLEDGMENTS
The authors would like to acknowledge KIST Medical College and Teaching Hospital for their support. Go to: Conflict of Interest
None. Go to: REFERENCES
Zeshan B, Karobari MI, Afzal N, Siddiq A, Basha S, Basheer SN, et al. The Usage of Antibiotics by COVID-19 Patients with Comorbidities: The Risk of Increased Antimicrobial Resistance. Antibiotics (Basel). 2021 Dec 29;11(1):35. doi: 10.3390/antibiotics11010035. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Rizvi SG, Ahammad SZ. COVID-19 and antimicrobial resistance: A cross-study. Sci Total Environ. 2022 Feb 10;807(Pt 2):150873. doi: 10.1016/j.scitotenv.2021.150873. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Chen N, Zhou M, Dong X, Qu J, Gong F, Han Y, et al. Epidemiological and clinical characteristics of 99 cases of 2019 novel coronavirus pneumonia in Wuhan, China: a descriptive study. Lancet. 2020 Feb 15;395(10223):507–13. doi: 10.1016/S0140-6736(20)30211-7. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Zhou F, Yu T, Du R, Fan G, Liu Y, Liu Z, et al. Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: a retrospective cohort study. Lancet. 2020 Mar 28;395(10229):1054–62. doi: 10.1016/S0140-6736(20)30566-3. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Molla MMA, Yeasmin M, Islam MK, Sharif MM, Amin MR, Nafisa T, et al. Antibiotic prescribing patterns at COVID-19 dedicated wards in Bangladesh: findings from a single center study. Infect Prev Pract. 2021 Jun;3(2):100134. doi: 10.1016/j.infpip.2021.100134. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Indian Council of Medical Research. Clinical guidance for management of adult COVID-19 patients [Internet]. New Delhi (IN): Indian Council of Medical Research; 2022. Jan 14, [2022 Jan 14; ]. [2022 Feb 20; ]. https://www.icmr.gov.in/ctechdocad.html Available from: [Google Scholar]
Barocas JA, Savinkina A, Lodi S, Epstein RL, Bouton TC, Sperring H, et al. Projected long-term impact of the COVID-19 pandemic on hepatitis C outcomes in the United States: a modelling study. Clin Infect Dis. 2021 Sep 9;:ciab779. doi: 10.1093/cid/ciab779. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Lucien MAB, Canarie MF, Kilgore PE, Jean-Denis G, Fenelon N, Pierre M, et al. Antibiotics and antimicrobial resistance in the COVID-19 era: Perspective from resource-limited settings. Int J Infect Dis. 2021 Mar;104:250–4. doi: 10.1016/j.ijid.2020.12.087. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Neto AGM, Lo KB, Wattoo A, Salacup G, Pelayo J, DeJoy R, et al. Bacterial infections and patterns of antibiotic use in patients with COVID-19. J Med Virol. 2021 Mar;93(3):1489–95. doi: 10.1002/jmv.26441. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Rawson TM, Moore LSP, Zhu N, Ranganathan N, Skolimowska K, Gilchrist M, et al. Bacterial and fungal coinfection in individuals with Coronavirus: A rapid review to support COVID-19 antimicrobial prescribing. Clin Infect Dis. 2020 Dec 3;71(9):2459–68. doi: 10.1093/cid/ciaa530. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Varghese GM, John R, Manesh A, Karthik R, Abraham OC. Clinical management of COVID-19. Indian J Med Res. 2020 May;151(5):401–10. doi: 10.4103/ijmr.IJMR_957_20. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Yacouba A, Olowo-Okere A, Yunusa I. Repurposing of antibiotics for clinical management of COVID-19: a narrative review. Ann Clin Microbiol Antimicrob. 2021 May 21;20(1):37. doi: 10.1186/s12941-021-00444-9. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Morens DM, Taubenberger JK, Fauci AS. Predominant role of bacterial pneumonia as a cause of death in pandemic influenza: implications for pandemic influenza preparedness. J Infect Dis. 2008 Oct 1;198(7):962–70. doi: 10.1086/591708. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Wei W, Ortwine JK, Mang NS, Joseph C, Hall BC, Prokesch BC. Limited Role for Antibiotics in COVID-19: Scarce evidence of bacterial coinfection. [2022 Feb 20; ];medRxiv [Preprint]. 2020 Jun 16; doi: 10.1101/2020.06.16.20133181. https://www.medrxiv.org/content/10.1101/2020.06.16.20133181v1 Available from: [CrossRef] [Google Scholar]
Mustafa L, Tolaj I, Baftiu N, Fejza H. Use of antibiotics in COVID-19 ICU patients. J Infect Dev Ctries. 2021 Apr 30;15(4):501–5. doi: 10.3855/jidc.14404. [PubMed] [CrossRef] [Google Scholar]
Grau S, Echeverria-Esnal D, Gomez-Zorrilla S, Navarrete-Rouco ME, Masclans JR, Espona M, et al. Evolution of antimicrobial consumption during the first wave of COVID-19 pandemic. Antibiotics (Basel). 2021 Jan 29;10(2):132. doi: 10.3390/antibiotics10020132. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Zhu N, Zhang D, Wang W, Li X, Yang B, Song J, et al. A Novel Coronavirus from patients with pneumonia in China, 2019. N Engl J Med. 2020 Feb 20;382(8):727–33. doi: 10.1056/NEJMoa2001017. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Morgan DJ, Casulli J, Chew C, Connolly E, Lui S, Brand OJ, Rahman R, Jagger C, Hussell T. innate immune cell suppression and the link with secondary lung bacterial pneumonia. Front Immunol. 2018 Dec 14;9:2943. doi: 10.3389/fimmu.2018.02943. [PMC free article] [PubMed] [CrossRef] [Google Scholar] THE PROBLEM ARISES FROM THE IMPACT OF A HEAVY DOSE OF ANTIBIOTICS ON THE MITOCHONDRIA AND ALSO THE LOSS OF IMMUNE SYSTEM FUNCTION COINCIDENT WITH LOSS OF MITOCHONDRIAL FUNCTION
Jyrkkanen, Jorma. (2020). Antibiotic induced changes to mitochondria result in potential contributions to carcinogenesis, heart pathologies, other medical conditions and ecosystem risks. Journal of Cardiology and Cardiovascular Medicine. 5. 163-171. 10.29328/journal.jccm.1001104.
In addition to providing energy to support the synthesis of the macromolecules essential for immune cell proliferation, mitochondria also act as signaling organelles, driving activation of immune cells via metabolic intermediates, mitochondrial DNA (mtDNA), and reactive oxygen species (ROS).Mar 22, 2022 Introduction
Immune dysregulation, characterized by an imbalance between a systemic inflammatory response syndrome and a compensatory anti-inflammatory response syndrome, is often observed in critically ill patients [1, 2]. This imbalance between the pro- and anti-inflammatory responses frequently leads to immunoparalysis in critically ill patients, rendering them more susceptible to further infections, and is associated with increased mortality [3]. Currently, no effective treatments are available to restore immune homeostasis and reduce mortality in these patients, largely due to the heterogeneity in patients’ immune status and more importantly the lack of understanding of the underlying cause of such immune dysfunction [2, 4]. Immune response is not a standalone process but is interconnected with other cellular activities, a very important one of which is cellular metabolism. Metabolic pathways and immune response are tightly intertwined both in health and in disease [5]. The link between immune cell function and mitochondrial function is now well recognized and a field known as “immunometabolism” is dedicated to understanding the relationship between immune and metabolic pathways [6,7,8]. Mitochondria play a crucial role in regulating not only the growth, but also the function, of immune cells. In addition to providing energy to support the synthesis of the macromolecules essential for immune cell proliferation, mitochondria also act as signaling organelles, driving activation of immune cells via metabolic intermediates, mitochondrial DNA (mtDNA), and reactive oxygen species (ROS). In addition, mitochondrial dynamics (fusion and fission), biogenesis (synthesis of new mitochondria), and mitophagy (degradation of damaged mitochondria) also play important roles in regulating immune cell functions. Knowledge in immunometabolism in critical illness, in particularly sepsis, opens up a new paradigm in patient care. Potential therapies targeting metabolic pathways, instead of solely immune-related pathways, might be the way to repair cellular function and restore immune homeostasis [4]. The other aspect of immunometabolism—looking at how immune responses influence metabolic pathways—is equally important, but beyond the scope of this review. Interaction between metabolism and immune response at the organ level has been reviewed elsewhere [6]. Mitochondrial Machinery That Mediates and Regulates Immune Responses in Critical Illness
Apart from being the powerhouse of the cell, the mitochondrion has emerged as a signaling hub that shapes and modulates how the immune system responds to infection or trauma. Mitochondrial dysfunction is evident in leukocytes from critically ill patients, and is believed to be the underlying cause of immunoparalysis and may account for the development of organ dysfunction [7,8,9]. Early recovery of mitochondrial function correlates with improved recovery in critically ill patients [10]. Metabolic Reprogramming
The immune-regulating mitochondrial machinery is a complex network involving many pathways and mechanisms that diverge and converge at various levels. Metabolic reprogramming is one mechanism that has been well studied in both innate and adaptive immune cells. Immune cells at different activation states (quiescent vs. activated), or with different functions (pro-inflammatory vs. anti-inflammatory), and different cell types (granulocytes, macrophages, dendritic cells, T- and B-lymphocytes), make use of different metabolic pathways (e.g., glycolysis, oxidative phosphorylation, fatty acid metabolism) to produce ATP [11]. The choice of different metabolic pathways, supports the energy demand of cells at different activation state. For example, upon infection or stimulation, immune cells become activated and produce cytokines and hence tend to favor glycolysis over oxidative phosphorylation for fast turnaround of ATP. Although the same amount of starting material, such as glucose, is used, oxidative phosphorylation generates 18 times more ATP than glycolysis, although is a lot slower. On the other hand, the choice of metabolic pathway determines the fate of the immune cells, i.e., naïve or memory, effector or regulatory, etc. However, the environment that the cells are in in the first place, triggers the changes in the metabolic pathways. The overall trend is that neutrophils, inflammatory macrophages (M1 macrophages), activated effector T cells, and dendritic cells rely more on aerobic glycolysis, whereas alternatively polarized macrophages (M2 macrophages), regulatory T cells (Tregs), and memory T cells prefer oxidative phosphorylation and fatty acid oxidation for energy production [8, 11, 12]. Metabolic reprogramming serves an important role in catering for the immune cells’ energy demand at different phases of their activation and proliferation. However, imbalance across the metabolic pathways could have serious pathological impact. One example may be the hyperlactatemia often seen in critically ill patients. Increased aerobic glycolysis in the activated immune cells during the initial hyper- inflammatory response is believed to contribute to the increase in blood lactate levels in sepsis [13, 14]. Mitochondrial ROS and mtDNA
Metabolic reprogramming sets the scene for the immune response, which is then subjected to many more modifications and regulations by factors that are directly or indirectly related to mitochondrial metabolism. Two important mitochondria-related immune regulators that have been well studied are mitochondrial ROS and mtDNA. Mitochondrial ROS are produced in healthy mitochondria, as a by-product of oxidative phosphorylation. At low dose, mitochondrial ROS serve important signaling functions, especially in the innate immune response. They are known to mediate NLRP3 inflammasome activation, leading to production of the pro-inflammatory cytokines, interleukin (IL)-1β and IL-18 [8, 15]. Mitochondrial ROS also induce a type-I interferon (IFN) response via mitochondrial antiviral-signaling (MAVS) and the IFN regulatory factor 3 (IRF3) pathway [16]. However, the level of mitochondrial ROS needs to be tightly regulated by the antioxidant system. Excessive mitochondrial ROS can cause oxidative damage to proteins/enzymes involved in oxidative phosphorylation and create mutations in mtDNA, contributing to the immune dysregulations as seen in critical illness [17]. Like mitochon-drial ROS, mtDNA also plays an important role in innate immunity [12]. In healthy cells, mtDNA is located in the matrix of mitochondria, encoding 13 proteins, all of which are components of oxidative phosphorylation. mtDNA is released to the cytosol upon mitochondrial dysfunction which involves changes to the integrity or permeability of the mitochondrial membrane. mtDNA, released into the cytosol, can activate the NLRP3 inflammasome with release of IL-1β and IL-18. Due to its bacterial origin, cytosolic mtDNA also serves as a damage-associated molecular pattern (DAMP), which can be recognized by intracellular pattern recognition receptors (PRRs), such as Toll-like receptor 9 (TLR9), and initiate the nuclear factor-kappa B (NF-κB)-dependent pro-inflammatory signaling pathway. In addition, cytosolic mtDNA can also be sensed by cyclic GMP-AMP synthase (cGAS) and activate the cGAS/stimulator of IFN genes (cGAS/STING) pathway and its downstream IFN response [18]. mtDNA can also be released into the circulation and cause systemic inflammation. Circulating mtDNA has been associated with mortality in critically ill patients [19]. Succinate and Itaconate
In addition to mitochondrial ROS and mtDNA, metabolites such as succinate and itaconate have also emerged as part of immune-regulating mitochondrial machinery [4, 20]. Both succinate and itaconate are intermediates from the tricarboxylic acid (TCA) cycle with opposite effects on the immune response. The TCA cycle generates nicotinamide adenine dinucleotide (NADH) and flavin adenine dinucleotide (FADH2), providing electrons to fuel oxidative phosphorylation. Succinate accumulation occurs under conditions such as hypoxia or inflammation. It can be released from mitochondria into the cytosol and functions as a signal transducer promoting pro-inflammatory gene expression via hypoxia-inducible factor 1α (HIF-1α) activation. Accumulation and oxidation of succinate by succinate dehydrogenase (SDH) in the mitochondria also leads to increased production of mitochondrial ROS via a process called reverse electron transport. This further enhances the pro- inflammatory effect of succinate. Like ROS, the level of succinate needs to be carefully regulated due to its inflammation aggravating effect. Plasma succinate has been proposed as a predictor of mortality for critically ill patients who are severely injured [21].
Itaconate, which is derived from cis-aconitate of the TCA cycle, is a succinate-regulating factor. It is shown to counteract the pro-inflammatory effect of succinate by inhibiting SDH. Itaconate can also be released into the cytosol and activate transcription factor NF-E2 p45-related factor 2 (Nrf2), a master regulator of antioxidant and anti-inflammatory responses [22]. Recently, itaconate has also been shown to inhibit the inflammatory response in macrophages through activating transcription factor 3 (ATF3). Mitochondrial Dynamics
The above mentioned immune-regulating mitochondrial factors are centered around the biochemical aspect of mitochondrial biology. Another important aspect of immune-regulating mitochondrial machinery is mitochondrial dynamics, which is to maintain and provide infrastructural support for the immune response. The size and shape of mitochondria undergo constant change through fusion and fission, which is important for maintaining the health and function of mitochondria. First, fusion incorporates newly synthesized mitochondria (from mitochondrial biogenesis) into the current mitochondrial network. Second, fusion also allows for mixing of proteins and/or mtDNA between the existing mitochondria, which on one hand enhances the metabolic capacity of the mitochondria, and on the other enables the damaged proteins and/or mutated mtDNA to be segregated from the healthy ones. Finally, segregation is achieved via fission and the damaged mitochondria can be destroyed through a process known as mitophagy. The proportion of mitochondria with damaged proteins or mutated mtDNA is kept below a critical threshold level through this process to maintain mitochondrial function [23, 24]. In addition to quality control, mitochondrial fusion and fission also participate in immune regulation. In activated T cells, there is an increase in fission, which creates round and fragmented mitochondria with loose cristae, favoring aerobic glycolysis. And in 146 memory T cells, increased fusion generates elongated mitochondria which favors oxidative phosphorylation and fatty acid oxidation [8, 25]. Immunometabolism: The Perfect World Scenario vs. the Critical Illness Scenario
So far, we have presented a list of mitochondrial components that are thought to play important roles in regulating the immune response. Our list is far from complete, but does highlight a few mechanisms that could relate to the development of immune dysregulation in critical illness. Figure 1 illustrates what we think would happen to the immune response when metabolism was in perfect control (the perfect world scenario) and when it became inconsistent and changeable (the critical illness scenario). In the perfect world scenario, the presence of an insult (e.g., infection or a trauma-related stress signal), would trigger metabolic reprogramming, switching from oxidative phosphorylation to glycolysis. This would enable activation of immune cells and production of pro-inflammatory cytokines and other mediators. At the same time, mitochondrial fission would increase to keep up with the metabolic reprogramming. The slightly elevated mitochondrial ROS and succinate in response to initial insult or cytokines would promote the pro-inflammatory response. Once the insult was eliminated, mitochondrial fusion would increase to create fused elongated mitochondria that favor oxidative phosphorylation and fatty acid oxidation. This would allow activation of regulatory immune cells and production of anti-inflammatory cytokines and other mediators. And itaconate would counteract the effect of succinate, activate the Nrf2-mediated antioxidant pathway to dampen down mitochondrial ROS, and activate ATF3 to inhibit the inflammatory response in macrophages. Immune homeostasis would be achieved as a result. Fig. 1 figure 1
Immunometabolism in the ‘perfect world scenario’ vs. the ‘critical illness scenario’. OXPHOS oxidative phosphorylation, FAO fatty acid oxidation Full size image
In the critical illness scenario, initial metabolic reprogramming from oxidative phosphorylation to glycolysis would go on for longer than necessary, generating excessive lactate (hyperlactatemia) and pro-inflammatory cytokines and mediators. A disrupted mitochondrial fusion/fission cycle could be to blame, one which could not support the timely switch to oxidative phosphorylation and fatty acid oxidation. The anti-inflammatory response would eventually kick in but by then damage would already have occurred to mitochondria and mtDNA because of excessive production of ROS in response to stress or cytokines. Excessive ROS and released mtDNA would aggravate the pro-inflammatory response, which in turn would trigger a more aggressive anti-inflammatory response to try and salvage the situation. The competition between pro- and anti-inflammatory responses would exhaust the nutrients and lead to shutdown of the whole metabolic system. Cells would either die or go into hibernation to preserve energy [26]. This scenario is an over-simplified version of what might happen in the actual disease setting, without considering the crosstalk between cells and organs and many other factors that are not included here. It is designed to shed light on the interaction between the immune response and metabolism. Potential of Mitochondria-Targeting Therapy in Critical Care
Our understanding thus far leads us to think that targeting mitochondria could perhaps correct the underlying cause of immune dysfunction in critical illness and lead to better recovery of the patients. The central role of mitochondrial dynamics in supporting and initiating metabolic reprogramming would make it the perfect therapeutic target. To get the fusion/fission cycle going, the mitochondrial network needs to be replenished by newly synthesized mitochondria via biogenesis. Therapies that could potentially boost mitochondrial biogenesis are mitochondrial transplantation, metformin, nitric oxide (NO), and carbon monoxide. Mitochondrial transplantation has been used successfully in pediatric patients with myocardial ischemia–reperfusion injury [27]. Metformin can activate peroxisome proliferator-activated receptor (PPAR)-gamma coactivator-1α (PGC-1α), and Nrf2, the master regulator of mitochondrial biogenesis and antioxidant systems [28]. Premorbid use of metfor-min is associated with lower mortality in sepsis [29]. NO and carbon monoxide can also enhance mitochondrial biogenesis [30,31,32]. Dietary nitrite has been trialed in patients with coronary artery disease (ClinicalTrials.gov Identifier: NCT00069654). Other therapies, such as mitochondria-targeted antioxidant (MitoQ) [33], could also be beneficial in protecting mtDNA and oxidative phosphorylation from oxidative damage. MitoQ has been trialed in people with Parkinson’s disease (ClinicalTrials. gov Identifier: NCT00329056). Challenges of Applying Mitochondria-Targeting Therapy in Critical Care
There are challenges to overcome before mitochondria-targeting therapy would be possible. First, how do we assess mitochondrial dysfunction in the clinic and identify patients who would benefit from such therapy? A few possible ways could be considered. Non-invasive assessment of mitochondrial oxygen metabolism using a novel device called the COMET monitor was tested on 40 patients during the acute phase of sepsis. This device is based on the protoporphyrin IX-triplet state lifetime technique (PpIX-TSLT) and has been shown to be feasible [33]. This technology is still in its early phase of clinical application but does offer some hope. Another possible biomarker that could potentially be used for assessing mitochondrial dysfunction is plasma mtDNA, but its sensitivity and specificity need further investigation [19, 34, 35]. Furthermore, we could consider using immune response markers as a surrogate markers, one such example could be IFNα inducible protein 27 (IFI27) [36]. If we could overcome the first challenge, the second would be how to deliver mitochondria-targeting therapies to the right organ at the right time. Conclusion
In this chapter, we have demonstrated the important role of mitochondria in regulating the immune response and proposed a scenario that explains immune–metabolism crosstalk in the context of critical illness. We have highlighted the role of mitochon-drial dynamics in overseeing and supporting metabolic reprogramming during immune cell activation. Mitochondrial ROS can be friend or foe when it comes to immune regulation. Two TCA intermediates—succinate and itaconate—with opposite effects have emerged as important players of the immune-regulating mitochon-drial machinery. Our understanding in immunometabolism could take us to the next era of critical care: mitochondria-targeting therapy. Availability of data and material
Not applicable. References
Duggal NA, Snelson C, Shaheen U, Pearce V, Lord JM. Innate and adaptive immune dysregulation in critically ill ICU patients. Sci Rep. 2018;8:10186.
Article
Google Scholar
Surbatovic M, Vojvodic D, Khan W. Immune response in critically ill patients. Mediat Inflamm. 2018;2018:9524315.
Article
Google Scholar
Frazier WJ, Hall MW. Immunoparalysis and adverse outcomes from critical illness. Pediatr Clin N Am. 2008;55:647–68.
Article
Google Scholar
Koutroulis I, Batabyal R, McNamara B, Ledda M, Hoptay C, Freishtat RJ. Sepsis immuno-metabolism: from defining sepsis to understanding how energy production affects immune response. Crit Care Explor. 2019;1:e0061.
Article
Google Scholar
Faas MM, de Vos P. Mitochondrial function in immune cells in health and disease. Biochim Biophys Acta Mol Basis Dis. 2020;1866:165845.
Article
CAS
Google Scholar
Lercher A, Baazim H, Bergthaler A. Systemic immunometabolism: challenges and opportunities. Immunity. 2020;53:496–509.
Article
CAS
Google Scholar
McBride MA, Owen AM, Stothers CL, et al. The metabolic basis of immune dysfunction following sepsis and trauma. Front Immunol. 2020;11:1043.
Article
CAS
Google Scholar
Angajala A, Lim S, Phillips JB, et al. Diverse roles of mitochondria in immune responses: novel insights into immuno-metabolism. Front Immunol. 2018;9:1605.
Article
Google Scholar
Cheng SC, Scicluna BP, Arts RJ, et al. Broad defects in the energy metabolism of leukocytes underlie immunoparalysis in sepsis. Nat Immunol. 2016;17:406–13.
Article
CAS
Google Scholar
Carré JE, Orban JC, Re L, et al. Survival in critical illness is associated with early activation of mitochondrial biogenesis. Am J Respir Crit Care Med. 2010;182:745–51.
Article
Google Scholar
Pearce EL, Pearce EJ. Metabolic pathways in immune cell activation and quiescence. Immunity. 2013;38:633–43.
Article
CAS
Google Scholar
Sack MN. Mitochondrial fidelity and metabolic agility control immune cell fate and function. J Clin Invest. 2018;128:3651–61.
Article
Google Scholar
Haji-Michael PG, Ladrière L, Sener A, Vincent JL, Malaisse WJ. Leukocyte glycolysis and lactate output in animal sepsis and ex vivo human blood. Metabolism. 1999;48:779–85.
Article
CAS
Google Scholar
Gibot S. On the origins of lactate during sepsis. Crit Care. 2012;16:151.
Article
Google Scholar
Zhou R, Yazdi AS, Menu P, Tschopp J. A role for mitochondria in NLRP3 inflammasome activation. Nature. 2011;469:221–5.
Article
CAS
Google Scholar
Agod Z, Fekete T, Budai MM, et al. Regulation of type I interferon responses by mitochondria-derived reactive oxygen species in plasmacytoid dendritic cells. Redox Biol. 2017;13:633–45.
Article
CAS
Google Scholar
Abilés J, de la Cruz AP, Castaño J, et al. Oxidative stress is increased in critically ill patients according to antioxidant vitamins intake, independent of severity: a cohort study. Crit Care. 2006;10:R146.
Article
Google Scholar
Riley JS, Tait SW. Mitochondrial DNA in inflammation and immunity. EMBO Rep. 2020;21:e49799.
Article
CAS
Google Scholar
Harrington JS, Huh JW, Schenck EJ, Nakahira K, Siempos II, Choi AMK. Circulating mitochondrial DNA as predictor of mortality in critically ill patients: a systematic review of clinical studies. Chest. 2019;156:1120–36.
Article
Google Scholar
Murphy MP, O’Neill LAJ. Krebs cycle reimagined: the emerging roles of succinate and itaconate as signal transducers. Cell. 2018;174:780–4.
Article
CAS
Google Scholar
D’Alessandro A, Moore HB, Moore EE, Reisz JA, Wither MJ, Ghasasbyan A, et al. Plasma succinate is a predictor of mortality in critically injured patients. J Trauma Acute Care Surg. 2017;83:491–5.
Article
Google Scholar
Mills EL, Ryan DG, Prag HA, et al. Itaconate is an anti-inflammatory metabolite that activates Nrf2 via alkylation of KEAP1. Nature. 2018;556:113–7.
Article
CAS
Google Scholar
Garbern JC, Lee RT. Mitochondria and metabolic transitions in cardiomyocytes: lessons from development for stem cell-derived cardiomyocytes. Stem Cell Res Ther. 2021;12:177.
Article
Google Scholar
Carelli V, Maresca A, Caporali L, Trifunov S, Zanna C, Rugolo M. Mitochondria: biogenesis and mitophagy balance in segregation and clonal expansion of mitochondrial DNA mutations. Int J Biochem Cell Biol. 2015;63:21–4.
Article
CAS
Google Scholar
Mills EL, Kelly B, O’Neill LAJ. Mitochondria are the powerhouses of immunity. Nat Immunol. 2017;18:488–98.
Article
CAS
Google Scholar
Singer M. The role of mitochondrial dysfunction in sepsis-induced multi-organ failure. Virulence. 2014;5:66–72.
Article
Google Scholar
McCully JD, Cowan DB, Emani SM, Del Nido PJ. Mitochondrial transplantation: from animal models to clinical use in humans. Mitochondrion. 2017;34:127–34.
Article
CAS
Google Scholar
Katila N, Bhurtel S, Park PH, Choi DY. Metformin attenuates rotenone-induced oxidative stress and mitochondrial damage via the AKT/Nrf2 pathway. Neurochem Int. 2021;148:105120.
Article
CAS
Google Scholar
Tan K, Simpson A, Huang S, Tang B, McLean A, Nalos M. The association of premorbid metformin exposure with mortality and organ dysfunction in sepsis: a systematic review and meta-analysis. Crit Care Explor. 2019;1:e0009.
Article
Google Scholar
Nisoli E, Clementi E, Paolucci C, et al. Mitochondrial biogenesis in mammals: the role of endogenous nitric oxide. Science. 2003;299:896–9.
Article
CAS
Google Scholar
Shiva S, Sack MN, Greer JJ, et al. Nitrite augments tolerance to ischemia/reperfusion injury via the modulation of mitochondrial electron transfer. J Exp Med. 2007;204:2089–102.
Article
CAS
Google Scholar
Lancel S, Hassoun SM, Favory R, Decoster B, Motterlini R, Neviere R. Carbon monoxide rescues mice from lethal sepsis by supporting mitochondrial energetic metabolism and activating mitochondrial biogenesis. J Pharmacol Exp Ther. 2009;329:641–8.
Article
CAS
Google Scholar
Lowes DA, Thottakam BM, Webster NR, Murphy MP, Galley HF. The mitochondria-targeted antioxidant MitoQ protects against organ damage in a lipopolysaccharide-peptidoglycan model of sepsis. Free Radic Biol Med. 2008;45:1559–65.
Article
CAS
Google Scholar
Faust HE, Reilly JP, Anderson BJ, et al. Plasma mitochondrial DNA levels are associated with ARDS in trauma and sepsis patients. Chest. 2020;157:67–76.
Article
CAS
Google Scholar
Mao JY, Li DK, Zhang HM, Wang XT, Liu DW. Plasma mitochondrial DNA levels are associated with acute lung injury and mortality in septic patients. BMC Pulm Med. 2021;21:66.
Article
CAS
Google Scholar
Tang BM, Shojaei M, Parnell GP, et al. A novel immune biomarker IFI27 discriminates between influenza and bacteria in patients with suspected respiratory infection. Eur Respir J. 2017;49:1602098.
The CEO of the OneAmerica insurance company publicly disclosed that during the third and fourth quarters of 2021, death in people of working age (18-64) was 40% higher than it was before the pandemic. Significantly, the majority of the deaths were not attributed to COVID. Even a… pic.twitter.com/Hx5EbBjgKn
2020, 2021,2022 Ed Dowd did Under an organization called the Humanities Project compile a history of excess deaths during and after the pandemic and here is what they found.
Question Did more all-cause deaths occur during the first months of the coronavirus disease 2019 (COVID-19) pandemic in the United States compared with the same months during previous years?
Findings 2020 In this cohort study, the number of deaths due to any cause increased by approximately 122 000 from March 1 to May 30, 2020, which is 28% higher than the reported number of COVID-19 deaths.
Meaning Official tallies of deaths due to COVID-19 underestimate the full increase in deaths associated with the pandemic in many states.
Abstract
Importance Efforts to track the severity and public health impact of coronavirus disease 2019 (COVID-19) in the United States have been hampered by state-level differences in diagnostic test availability, differing strategies for prioritization of individuals for testing, and delays between testing and reporting. Evaluating unexplained increases in deaths due to all causes or attributed to nonspecific outcomes, such as pneumonia and influenza, can provide a more complete picture of the burden of COVID-19.
Objective To estimate the burden of all deaths related to COVID-19 in the United States from March to May 2020.
Design, Setting, and Population This observational study evaluated the numbers of US deaths from any cause and deaths from pneumonia, influenza, and/or COVID-19 from March 1 through May 30, 2020, using public data of the entire US population from the National Center for Health Statistics (NCHS). These numbers were compared with those from the same period of previous years. All data analyzed were accessed on June 12, 2020.
Main Outcomes and Measures Increases in weekly deaths due to any cause or deaths due to pneumonia/influenza/COVID-19 above a baseline, which was adjusted for time of year, influenza activity, and reporting delays. These estimates were compared with reported deaths attributed to COVID-19 and with testing data.
Results There were approximately 781 000 total deaths in the United States from March 1 to May 30, 2020, representing 122 300 (95% prediction interval, 116 800-127 000) more deaths than would typically be expected at that time of year. There were 95 235 reported deaths officially attributed to COVID-19 from March 1 to May 30, 2020. The number of excess all-cause deaths was 28% higher than the official tally of COVID-19–reported deaths during that period. In several states, these deaths occurred before increases in the availability of COVID-19 diagnostic tests and were not counted in official COVID-19 death records. There was substantial variability between states in the difference between official COVID-19 deaths and the estimated burden of excess deaths.
Conclusions and Relevance Excess deaths provide an estimate of the full COVID-19 burden and indicate that official tallies likely undercount deaths due to the virus. The mortality burden and the completeness of the tallies vary markedly between states.
Discussion
Monitoring excess deaths has been used as a method for tracking influenza mortality for more than a century. Herein, we used a similar strategy to capture COVID-19 deaths that had not been attributed specifically to the pandemic coronavirus. Monitoring trends in broad mortality outcomes, like changes in all-cause and pneumonia/influenza/COVID-19 mortality, provides a window into the magnitude of the mortality burden missed in official tallies of COVID-19 deaths. Given the variability in testing intensity between states and over time, this type of monitoring provides key information on the severity of the pandemic and the degree to which viral testing might be missing deaths caused by COVID-19. These findings demonstrate that estimates of the death toll of COVID-19 based on excess all-cause mortality may be more reliable than those relying only on reported deaths, particularly in places that lack widespread testing.
Syndromic end points, such as deaths due to pneumonia/influenza/COVID-19, outpatient visits for influenza-like illness, and emergency department visits for fever, can provide a crude but informative measure of the progression of the outbreak.18 These measures themselves can be biased by changes in health-seeking behavior and how conditions are recorded. However, in the absence of widespread and systematic testing for COVID-19, they provide a useful measure of pandemic progression and the impact of interventions.
The gap between reported COVID-19 deaths and excess deaths can be influenced by several factors, including the intensity of testing; guidelines on the recording of deaths that are suspected to be related to COVID-19 but do not have a laboratory confirmation; and the location of death (eg, hospital, nursing home, or unattended death at home). For instance, deaths that occur in nursing homes might be more likely to be recognized as part of an epidemic and correctly recorded as due to COVID-19. As the pandemic has progressed, official statistics have become better aligned with excess mortality estimates, perhaps due to enhanced testing and increased recognition of the clinical features of COVID-19. In New York City, official COVID-19 death counts were revised after careful inspection of death certificates, adding an extra 5048 probable deaths to the 13 831 laboratory-confirmed deaths.19 As a result, the all-cause excess mortality burden from March 11 to May 2, 2020, is only 27% higher than official COVID-19 statistics.19 This aligns well with our estimate of 26% for a similar period in New York City, using a slightly different modeling approach.
Many European countries have experienced sharp increases in all-cause deaths associated with the pandemic. Real-time all-cause mortality data from the EuroMomo project (https://www.euromomo.eu/) demonstrate gaps between the official COVID-19 death toll and excess deaths that echo findings in our study. These gaps are more pronounced in countries that were affected more and earlier by the pandemic and had weak testing. Very limited excess mortality information is available from Asia, Africa, the Middle East, and South America thus far; these data will be important to fully capture the heterogeneity of death rates related to the COVID-19 pandemic across the world. Prior work on the 1918 and 2009 pandemics has shown substantial heterogeneity in mortality burden between countries, in part related to health care.8,20
Humanities Project Excess Deaths USA by Year and Vaccination Status
Horowitz Possible Explanation from Pfizer Databank
A SARS-like cluster of circulating bat coronaviruses shows potential for human emergence and study indicate that group 2b viruses encoding the SHC014 spike in a wild-type backbone can efficiently use multiple orthologs of the SARS receptor human angiotensin converting enzyme II (ACE2), replicate efficiently in primary human airway cells and achieve in vitro titers equivalent to epidemic strains of SARS-CoV. Repost w emphasis on spike used by Baric and Daszak.
Associated Revelation. Wuhan Institute of Virology Shao Cao admits they Tested Coronavirus Variants for the Best Adeherent to ACE2 Receptor
Abstract
The emergence of severe acute respiratory syndrome coronavirus (SARS-CoV) and Middle East respiratory syndrome (MERS)-CoV underscores the threat of cross-species transmission events leading to outbreaks in humans. Here we examine the disease potential of a SARS-like virus, SHC014-CoV, which is currently circulating in Chinese horseshoe bat populations1. Using the SARS-CoV reverse genetics system2, we generated and characterized a chimeric virus expressing the spike of bat coronavirus SHC014 in a mouse-adapted SARS-CoV backbone. The results indicate that group 2b viruses encoding the SHC014 spike in a wild-type backbone can efficiently use multiple orthologs of the SARS receptor human angiotensin converting enzyme II (ACE2), replicate efficiently in primary human airway cells and achieve in vitro titers equivalent to epidemic strains of SARS-CoV. Additionally, in vivo experiments demonstrate replication of the chimeric virus in mouse lung with notable pathogenesis. Evaluation of available SARS-based immune-therapeutic and prophylactic modalities revealed poor efficacy; both monoclonal antibody and vaccine approaches failed to neutralize and protect from infection with CoVs using the novel spike protein. On the basis of these findings, we synthetically re-derived an infectious full-length SHC014 recombinant virus and demonstrate robust viral replication both in vitro and in vivo. Our work suggests a potential risk of SARS-CoV re-emergence from viruses currently circulating in bat populations.
Supplementary information
The online version of this article (doi:10.1038/nm.3985) contains supplementary material, which is available to authorized users.
Subject terms: Policy and public health in microbiology, Viral infection, SARS virus, Translational research
SPIKE PROTEIN CLOTTING PROBLEM
Dr. McCullough: The COVID Vaccines Are Causing the ‘Largest Blood Clots We’ve Ever Seen’
“My experience with these blood clots are they’re enormously resistant to blood thinners,” disclosed Dr. @P_McCulloughMD. “Wuhan spike protein is inside the blood clots, and it’s folding,… pic.twitter.com/UIawzXBToH
The emergence of SARS-CoV heralded a new era in the cross-species transmission of severe respiratory illness with globalization leading to rapid spread around the world and massive economic impact3,4. Since then, several strains—including influenza A strains H5N1, H1N1 and H7N9 and MERS-CoV—have emerged from animal populations, causing considerable disease, mortality and economic hardship for the afflicted regions5. Although public health measures were able to stop the SARS-CoV outbreak4, recent metagenomics studies have identified sequences of closely related SARS-like viruses circulating in Chinese bat populations that may pose a future threat1,6. However, sequence data alone provides minimal insights to identify and prepare for future prepandemic viruses. Therefore, to examine the emergence potential (that is, the potential to infect humans) of circulating bat CoVs, we built a chimeric virus encoding a novel, zoonotic CoV spike protein—from the RsSHC014-CoV sequence that was isolated from Chinese horseshoe bats1—in the context of the SARS-CoV mouse-adapted backbone. The hybrid virus allowed us to evaluate the ability of the novel spike protein to cause disease independently of other necessary adaptive mutations in its natural backbone. Using this approach, we characterized CoV infection mediated by the SHC014 spike protein in primary human airway cells and in vivo, and tested the efficacy of available immune therapeutics against SHC014-CoV. Together, the strategy translates metagenomics data to help predict and prepare for future emergent viruses.
The sequences of SHC014 and the related RsWIV1-CoV show that these CoVs are the closest relatives to the epidemic SARS-CoV strains (Fig. 1a,b); however, there are important differences in the 14 residues that bind human ACE2, the receptor for SARS-CoV, including the five that are critical for host range: Y442, L472, N479, T487 and Y491 (ref. 7). In WIV1, three of these residues vary from the epidemic SARS-CoV Urbani strain, but they were not expected to alter binding to ACE2 (Supplementary Fig. 1a,b and Supplementary Table 1). This fact is confirmed by both pseudotyping experiments that measured the ability of lentiviruses encoding WIV1 spike proteins to enter cells expressing human ACE2 (Supplementary Fig. 1) and by in vitro replication assays of WIV1-CoV (ref. 1). In contrast, 7 of 14 ACE2-interaction residues in SHC014 are different from those in SARS-CoV, including all five residues critical for host range (Supplementary Fig. 1c and Supplementary Table 1). These changes, coupled with the failure of pseudotyped lentiviruses expressing the SHC014 spike to enter cells (Supplementary Fig. 1d), suggested that the SHC014 spike is unable to bind human ACE2. However, similar changes in related SARS-CoV strains had been reported to allow ACE2 binding7,8, suggesting that additional functional testing was required for verification. Therefore, we synthesized the SHC014 spike in the context of the replication-competent, mouse-adapted SARS-CoV backbone (we hereafter refer to the chimeric CoV as SHC014-MA15) to maximize the opportunity for pathogenesis and vaccine studies in mice (Supplementary Fig. 2a). Despite predictions from both structure-based modeling and pseudotyping experiments, SHC014-MA15 was viable and replicated to high titers in Vero cells (Supplementary Fig. 2b). Similarly to SARS, SHC014-MA15 also required a functional ACE2 molecule for entry and could use human, civet and bat ACE2 orthologs (Supplementary Fig. 2c,d). To test the ability of the SHC014 spike to mediate infection of the human airway, we examined the sensitivity of the human epithelial airway cell line Calu-3 2B4 (ref. 9) to infection and found robust SHC014-MA15 replication, comparable to that of SARS-CoV Urbani (Fig. 1c). To extend these findings, primary human airway epithelial (HAE) cultures were infected and showed robust replication of both viruses (Fig. 1d). Together, the data confirm the ability of viruses with the SHC014 spike to infect human airway cells and underscore the potential threat of cross-species transmission of SHC014-CoV.
SARS-like viruses replicate in human airway cells and produce in vivo pathogenesis.
(a) The full-length genome sequences of representative CoVs were aligned and phylogenetically mapped as described in the Online Methods. The scale bar represents nucleotide substitutions, with only bootstrap support above 70% being labeled. The tree shows CoVs divided into three distinct phylogenetic groups, defined as α-CoVs, β-CoVs and γ-CoVs. Classical subgroup clusters are marked as 2a, 2b, 2c and 2d for the β-CoVs and as 1a and 1b for the α-CoVs. (b) Amino acid sequences of the S1 domains of the spikes of representative β-CoVs of the 2b group, including SARS-CoV, were aligned and phylogenetically mapped. The scale bar represents amino acid substitutions. (c,d) Viral replication of SARS-CoV Urbani (black) and SHC014-MA15 (green) after infection of Calu-3 2B4 cells (c) or well-differentiated, primary air-liquid interface HAE cell cultures (d) at a multiplicity of infection (MOI) of 0.01 for both cell types. Samples were collected at individual time points with biological replicates (n = 3) for both Calu-3 and HAE experiments. (e,f) Weight loss (n = 9 for SARS-CoV MA15; n = 16 for SHC014-MA15) (e) and viral replication in the lungs (n = 3 for SARS-CoV MA15; n = 4 for SHC014-MA15) (f) of 10-week-old BALB/c mice infected with 1 × 104 p.f.u. of mouse-adapted SARS-CoV MA15 (black) or SHC014-MA15 (green) via the intranasal (i.n.) route. (g,h) Representative images of lung sections stained for SARS-CoV N antigen from mice infected with SARS-CoV MA15 (n = 3 mice) (g) or SHC014-MA15 (n = 4 mice) (h) are shown. For each graph, the center value represents the group mean, and the error bars define the s.e.m. Scale bars, 1 mm.
To evaluate the role of the SHC014 spike in mediating infection in vivo, we infected 10-week-old BALB/c mice with 104 plaque-forming units (p.f.u.) of either SARS-MA15 or SHC014-MA15 (Fig. 1e–h). Animals infected with SARS-MA15 experienced rapid weight loss and lethality by 4 d post infection (d.p.i.); in contrast, SHC014-MA15 infection produced substantial weight loss (10%) but no lethality in mice (Fig. 1e). Examination of viral replication revealed nearly equivalent viral titers from the lungs of mice infected with SARS-MA15 or SHC014-MA15 (Fig. 1f). Whereas lungs from the SARS-MA15–infected mice showed robust staining in both the terminal bronchioles and the lung parenchyma 2 d.p.i. (Fig. 1g), those of SHC014-MA15–infected mice showed reduced airway antigen staining (Fig. 1h); in contrast, no deficit in antigen staining was observed in the parenchyma or in the overall histology scoring, suggesting differential infection of lung tissue for SHC014-MA15 (Supplementary Table 2). We next analyzed infection in more susceptible, aged (12-month-old) animals. SARS-MA15–infected animals rapidly lost weight and succumbed to infection (Supplementary Fig. 3a,b). SHC014-MA15 infection induced robust and sustained weight loss, but had minimal lethality. Trends in the histology and antigen staining patterns that we observed in young mice were conserved in the older animals (Supplementary Table 3). We excluded the possibility that SHC014-MA15 was mediating infection through an alternative receptor on the basis of experiments using Ace2−/− mice, which did not show weight loss or antigen staining after SHC014-MA15 infection (Supplementary Fig. 4a,b and Supplementary Table 2). Together, the data indicate that viruses with the SHC014 spike are capable of inducing weight loss in mice in the context of a virulent CoV backbone.
Given the preclinical efficacy of Ebola monoclonal antibody therapies, such as ZMApp10, we next sought to determine the efficacy of SARS-CoV monoclonal antibodies against infection with SHC014-MA15. Four broadly neutralizing human monoclonal antibodies targeting SARS-CoV spike protein had been previously reported and are probable reagents for immunotherapy11,12,13. We examined the effect of these antibodies on viral replication (expressed as percentage inhibition of viral replication) and found that whereas wild-type SARS-CoV Urbani was strongly neutralized by all four antibodies at relatively low antibody concentrations (Fig. 2a–d), neutralization varied for SHC014-MA15. Fm6, an antibody generated by phage display and escape mutants11,12, achieved only background levels of inhibition of SHC014-MA15 replication (Fig. 2a). Similarly, antibodies 230.15 and 227.14, which were derived from memory B cells of SARS-CoV–infected patients13, also failed to block SHC014-MA15 replication (Fig. 2b,c). For all three antibodies, differences between the SARS and SHC014 spike amino acid sequences corresponded to direct or adjacent residue changes found in SARS-CoV escape mutants (fm6 N479R; 230.15 L443V; 227.14 K390Q/E), which probably explains the absence of the antibodies’ neutralizing activity against SHC014. Finally, monoclonal antibody 109.8 was able to achieve 50% neutralization of SHC014-MA15, but only at high concentrations (10 μg/ml) (Fig. 2d). Together, the results demonstrate that broadly neutralizing antibodies against SARS-CoV may only have marginal efficacy against emergent SARS-like CoV strains such as SHC014.
SARS-CoV monoclonal antibodies have marginal efficacy against SARS-like CoVs.
(a–d) Neutralization assays evaluating efficacy (measured as reduction in the number of plaques) of a panel of monoclonal antibodies, which were all originally generated against epidemic SARS-CoV, against infection of Vero cells with SARS-CoV Urbani (black) or SHC014-MA15 (green). The antibodies tested were fm6 (n = 3 for Urbani; n = 5 for SHC014-MA15)11,12 (a), 230.15 (n = 3 for Urbani; n = 2 for SHC014-MA15) (b), 227.15 (n = 3 for Urbani; n = 5 for SHC014-MA15) (c) and 109.8 (n = 3 for Urbani; n = 2 for SHC014-MA15)13 (d). Each data point represents the group mean and error bars define the s.e.m. Note that the error bars in SARS-CoV Urbani–infected Vero cells in b,c are overlapped by the symbols and are not visible.
To evaluate the efficacy of existing vaccines against infection with SHC014-MA15, we vaccinated aged mice with double-inactivated whole SARS-CoV (DIV). Previous work showed that DIV could neutralize and protect young mice from challenge with a homologous virus14; however, the vaccine failed to protect aged animals in which augmented immune pathology was also observed, indicating the possibility of the animals being harmed because of the vaccination15. Here we found that DIV did not provide protection from challenge with SHC014-MA15 with regards to weight loss or viral titer (Supplementary Fig. 5a,b). Consistent with a previous report with other heterologous group 2b CoVs15, serum from DIV-vaccinated, aged mice also failed to neutralize SHC014-MA15 (Supplementary Fig. 5c). Notably, DIV vaccination resulted in robust immune pathology (Supplementary Table 4) and eosinophilia (Supplementary Fig. 5d–f). Together, these results confirm that the DIV vaccine would not be protective against infection with SHC014 and could possibly augment disease in the aged vaccinated group.
In contrast to vaccination of mice with DIV, the use of SHC014-MA15 as a live, attenuated vaccine showed potential cross-protection against challenge with SARS-CoV, but the results have important caveats. We infected young mice with 104 p.f.u. of SHC014-MA15 and observed them for 28 d. We then challenged the mice with SARS-MA15 at day 29 (Supplementary Fig. 6a). The prior infection of the mice with the high dose of SHC014-MA15 conferred protection against challenge with a lethal dose of SARS-MA15, although there was only a minimal SARS-CoV neutralization response from the antisera elicited 28 d after SHC014-MA15 infection (Supplementary Fig. 6b, 1:200). In the absence of a secondary antigen boost, 28 d.p.i. represents the expected peak of antibody titers and implies that there will be diminished protection against SARS-CoV over time16,17. Similar results showing protection against challenge with a lethal dose of SARS-CoV were observed in aged BALB/c mice with respect to weight loss and viral replication (Supplementary Fig. 6c,d). However, the SHC014-MA15 infection dose of 104 p.f.u. induced >10% weight loss and lethality in some aged animals (Fig. 1 and Supplementary Fig. 3). We found that vaccination with a lower dose of SHC014-MA15 (100 p.f.u.), did not induce weight loss, but it also failed to protect aged animals from a SARS-MA15 lethal dose challenge (Supplementary Fig. 6e,f). Together, the data suggest that SHC014-MA15 challenge may confer cross-protection against SARS-CoV through conserved epitopes, but the required dose induces pathogenesis and precludes use as an attenuated vaccine.
Having established that the SHC014 spike has the ability to mediate infection of human cells and cause disease in mice, we next synthesized a full-length SHC014-CoV infectious clone based on the approach used for SARS-CoV (Fig. 3a)2. Replication in Vero cells revealed no deficit for SHC014-CoV relative to that for SARS-CoV (Fig. 3b); however, SHC014-CoV was significantly (P < 0.01) attenuated in primary HAE cultures at both 24 and 48 h after infection (Fig. 3c). In vivo infection of mice demonstrated no significant weight loss but showed reduced viral replication in lungs of full-length SHC014-CoV infection, as compared to SARS-CoV Urbani (Fig. 3d,e). Together, the results establish the viability of full-length SHC014-CoV, but suggest that further adaptation is required for its replication to be equivalent to that of epidemic SARS-CoV in human respiratory cells and in mice.
Full-length SHC014-CoV replicates in human airways but lacks the virulence of epidemic SARS-CoV.
(a) Schematic of the SHC014-CoV molecular clone, which was synthesized as six contiguous cDNAs (designated SHC014A, SHC014B, SHC014C, SHC014D, SHC014E and SHC014F) flanked by unique BglI sites that allowed for directed assembly of the full-length cDNA expressing open reading frames (for 1a, 1b, spike, 3, envelope, matrix, 6–8 and nucleocapsid). Underlined nucleotides represent the overhang sequences formed after restriction enzyme cleavage. (b,c) Viral replication of SARS-CoV Urbani (black) or SHC014-CoV (green) after infection of Vero cells (b) or well-differentiated, primary air-liquid interface HAE cell cultures (c) at an MOI of 0.01. Samples were collected at individual time points with biological replicates (n = 3) for each group. Data represent one experiment for both Vero and HAE cells. (d,e) Weight loss (n = 3 for SARS-CoV MA15, n = 7 for SHC014-CoV; n = 6 for SARS-Urbani) (d) and viral replication in the lungs (n = 3 for SARS-Urbani and SHC014-CoV) (e) of 10-week-old BALB/c mice infected with 1 × 105 p.f.u. of SARS-CoV MA15 (gray), SHC014-CoV (green) or SARS-CoV Urbani (black) via the i.n. route. Each data point represents the group mean, and error bars define the s.e.m. **P < 0.01 and ***P < 0.001 using two-tailed Student’s t-test of individual time points.
During the SARS-CoV epidemic, links were quickly established between palm civets and the CoV strains that were detected in humans4. Building on this finding, the common emergence paradigm argues that epidemic SARS-CoV originated as a bat virus, jumped to civets and incorporated changes within the receptor-binding domain (RBD) to improve binding to civet Ace2 (ref. 18). Subsequent exposure to people in live-animal markets permitted human infection with the civet strain, which, in turn, adapted to become the epidemic strain (Fig. 4a). However, phylogenetic analysis suggests that early human SARS strains appear more closely related to bat strains than to civet strains18. Therefore, a second paradigm argues that direct bat-human transmission initiated SARS-CoV emergence and that palm civets served as a secondary host and reservoir for continued infection (Fig. 4b)19. For both paradigms, spike adaptation in a secondary host is seen as a necessity, with most mutations expected to occur within the RBD, thereby facilitating improved infection. Both theories imply that pools of bat CoVs are limited and that host-range mutations are both random and rare, reducing the likelihood of future emergence events in humans.
Coronavirus strains are maintained in quasi-species pools circulating in bat populations. (a,b) Traditional SARS-CoV emergence theories posit that host-range mutants (red circle) represent random and rare occurrences that permit infection of alternative hosts. The secondary-host paradigm (a) argues that a nonhuman host is infected by a bat progenitor virus and, through adaptation, facilitates transmission to humans; subsequent replication in humans leads to the epidemic viral strain. The direct paradigm (b) suggests that transmission occurs between bats and humans without the requirement of an intermediate host; selection then occurs in the human population with closely related viruses replicating in a secondary host, permitting continued viral persistence and adaptation in both. (c) The data from chimeric SARS-like viruses argue that the quasi-species pools maintain multiple viruses capable of infecting human cells without the need for mutations (red circles). Although adaptations in secondary or human hosts may be required for epidemic emergence, if SHC014 spike–containing viruses recombined with virulent CoV backbones (circles with green outlines), then epidemic disease may be the result in humans. Existing data support elements of all three paradigms.
Although our study does not invalidate the other emergence routes, it does argue for a third paradigm in which circulating bat CoV pools maintain ‘poised’ spike proteins that are capable of infecting humans without mutation or adaptation (Fig. 4c). This hypothesis is illustrated by the ability of a chimeric virus containing the SHC014 spike in a SARS-CoV backbone to cause robust infection in both human airway cultures and in mice without RBD adaptation. Coupled with the observation of previously identified pathogenic CoV backbones3,20, our results suggest that the starting materials required for SARS-like emergent strains are currently circulating in animal reservoirs. Notably, although full-length SHC014-CoV probably requires additional backbone adaption to mediate human disease, the documented high-frequency recombination events in CoV families underscores the possibility of future emergence and the need for further preparation.
To date, genomics screens of animal populations have primarily been used to identify novel viruses in outbreak settings21. The approach here extends these data sets to examine questions of viral emergence and therapeutic efficacy. We consider viruses with the SHC014 spike a potential threat owing to their ability to replicate in primary human airway cultures, the best available model for human disease. In addition, the observed pathogenesis in mice indicates a capacity for SHC014-containing viruses to cause disease in mammalian models, without RBD adaptation. Notably, differential tropism in the lung as compared to that with SARS-MA15 and attenuation of full-length SHC014-CoV in HAE cultures relative to SARS-CoV Urbani suggest that factors beyond ACE2 binding—including spike processivity, receptor bio-availability or antagonism of the host immune responses—may contribute to emergence. However, further testing in nonhuman primates is required to translate these finding into pathogenic potential in humans. Importantly, the failure of available therapeutics defines a critical need for further study and for the development of treatments. With this knowledge, surveillance programs, diagnostic reagents and effective treatments can be produced that are protective against the emergence of group 2b–specific CoVs, such as SHC014, and these can be applied to other CoV branches that maintain similarly heterogeneous pools.
In addition to offering preparation against future emerging viruses, this approach must be considered in the context of the US government–mandated pause on gain-of-function (GOF) studies22. On the basis of previous models of emergence (Fig. 4a,b), the creation of chimeric viruses such as SHC014-MA15 was not expected to increase pathogenicity. Although SHC014-MA15 is attenuated relative to its parental mouse-adapted SARS-CoV, similar studies examining the pathogenicity of CoVs with the wild-type Urbani spike within the MA15 backbone showed no weight loss in mice and reduced viral replication23. Thus, relative to the Urbani spike–MA15 CoV, SHC014-MA15 shows a gain in pathogenesis (Fig. 1). On the basis of these findings, scientific review panels may deem similar studies building chimeric viruses based on circulating strains too risky to pursue, as increased pathogenicity in mammalian models cannot be excluded. Coupled with restrictions on mouse-adapted strains and the development of monoclonal antibodies using escape mutants, research into CoV emergence and therapeutic efficacy may be severely limited moving forward. Together, these data and restrictions represent a crossroads of GOF research concerns; the potential to prepare for and mitigate future outbreaks must be weighed against the risk of creating more dangerous pathogens. In developing policies moving forward, it is important to consider the value of the data generated by these studies and whether these types of chimeric virus studies warrant further investigation versus the inherent risks involved.
Overall, our approach has used metagenomics data to identify a potential threat posed by the circulating bat SARS-like CoV SHC014. Because of the ability of chimeric SHC014 viruses to replicate in human airway cultures, cause pathogenesis in vivo and escape current therapeutics, there is a need for both surveillance and improved therapeutics against circulating SARS-like viruses. Our approach also unlocks the use of metagenomics data to predict viral emergence and to apply this knowledge in preparing to treat future emerging virus infections.
Viruses, cells, in vitro infection and plaque assays.
Wild-type SARS-CoV (Urbani), mouse-adapted SARS-CoV (MA15) and chimeric SARS-like CoVs were cultured on Vero E6 cells (obtained from United States Army Medical Research Institute of Infectious Diseases), grown in Dulbecco’s modified Eagle’s medium (DMEM) (Gibco, CA) and 5% fetal clone serum (FCS) (Hyclone, South Logan, UT) along with antibiotic/antimycotic (Gibco, Carlsbad, CA). DBT cells (Baric laboratory, source unknown) expressing ACE2 orthologs have been previously described for both human and civet; bat Ace2 sequence was based on that from Rhinolophus leschenaulti, and DBT cells expressing bat Ace2 were established as described previously8. Pseudotyping experiments were similar to those using an HIV-based pseudovirus, prepared as previously described10, and examined on HeLa cells (Wuhan Institute of Virology) that expressed ACE2 orthologs. HeLa cells were grown in minimal essential medium (MEM) (Gibco, CA) supplemented with 10% FCS (Gibco, CA) as previously described24. Growth curves in Vero E6, DBT, Calu-3 2B4 and primary human airway epithelial cells were performed as previously described8,25. None of the working cell line stocks were authenticated or tested for mycoplasma recently, although the original seed stocks used to create the working stocks are free from contamination. Human lungs for HAE cultures were procured under University of North Carolina at Chapel Hill Institutional Review Board–approved protocols. HAE cultures represent highly differentiated human airway epithelium containing ciliated and non-ciliated epithelial cells as well as goblet cells. The cultures are also grown on an air-liquid interface for several weeks before use, as previously described26. Briefly, cells were washed with PBS and inoculated with virus or mock-diluted in PBS for 40 min at 37 °C. After inoculation, cells were washed three times and fresh medium was added to signify time ‘0’. Three or more biological replicates were harvested at each described time point. No blinding was used in any sample collections nor were samples randomized. All virus cultivation was performed in a biosafety level (BSL) 3 laboratory with redundant fans in the biosafety cabinets, as described previously by our group2. All personnel wore powered air purifying respirators (Breathe Easy, 3M) with Tyvek suits, aprons and booties and were double-gloved.
Sequence clustering and structural modeling.
The full-length genomic sequences and the amino acid sequences of the S1 domains of the spike of representative CoVs were downloaded from Genbank or Pathosystems Resource Integration Center (PATRIC), aligned with ClustalX and phylogenetically compared by using maximum likelihood estimation using 100 bootstraps or by using the PhyML (https://code.google.com/p/phyml/) package, respectively. The tree was generated using maximum likelihood with the PhyML package. The scale bar represents nucleotide substitutions. Only nodes with bootstrap support above 70% are labeled. The tree shows that CoVs are divided into three distinct phylogenetic groups defined as α-CoVs, β-CoVs and γ-CoVs. Classical subgroup clusters are marked as 2a, 2b, 2c and 2d for β-CoVs, and 1a and 1b for the α-CoVs. Structural models were generated using Modeller (Max Planck Institute Bioinformatics Toolkit) to generate homology models for SHC014 and Rs3367 of the SARS RBD in complex with ACE2 based on crystal structure 2AJF (Protein Data Bank). Homology models were visualized and manipulated in MacPyMol (version 1.3).
Construction of SARS-like chimeric viruses.
Both wild-type and chimeric viruses were derived from either SARS-CoV Urbani or the corresponding mouse-adapted (SARS-CoV MA15) infectious clone (ic) as previously described27. Plasmids containing spike sequences for SHC014 were extracted by restriction digest and ligated into the E and F plasmid of the MA15 infectious clone. The clone was designed and purchased from Bio Basic as six contiguous cDNAs using published sequences flanked by unique class II restriction endonuclease sites (BglI). Thereafter, plasmids containing wild-type, chimeric SARS-CoV and SHC014-CoV genome fragments were amplified, excised, ligated and purified. In vitro transcription reactions were then preformed to synthesize full-length genomic RNA, which was transfected into Vero E6 cells as previously described2. The medium from transfected cells was harvested and served as seed stocks for subsequent experiments. Chimeric and full-length viruses were confirmed by sequence analysis before use in these studies. Synthetic construction of chimeric mutant and full-length SHC014-CoV was approved by the University of North Carolina Institutional Biosafety Committee and the Dual Use Research of Concern committee.
Ethics statement.
This study was carried out in accordance with the recommendations for the care and use of animals by the Office of Laboratory Animal Welfare (OLAW), NIH. The Institutional Animal Care and Use Committee (IACUC) of The University of North Carolina at Chapel Hill (UNC, Permit Number A-3410-01) approved the animal study protocol (IACUC #13-033) used in these studies.
Mice and in vivo infection.
Female, 10-week-old and 12-month-old BALB/cAnNHsD mice were ordered from Harlan Laboratories. Mouse infections were done as previously described20. Briefly, animals were brought into a BSL3 laboratory and allowed to acclimate for 1 week before infection. For infection and live-attenuated virus vaccination, mice were anesthetized with a mixture of ketamine and xylazine and infected intranasally, when challenged, with 50 μl of phosphate-buffered saline (PBS) or diluted virus with three or four mice per time point, per infection group per dose as described in the figure legends. For individual mice, notations for infection including failure to inhale the entire dose, bubbling of inoculum from the nose, or infection through the mouth may have led to exclusion of mouse data at the discretion of the researcher; post-infection, no other pre-established exclusion or inclusion criteria are defined. No blinding was used in any animal experiments, and animals were not randomized. For vaccination, young and aged mice were vaccinated by footpad injection with a 20-μl volume of either 0.2 μg of double-inactivated SARS-CoV vaccine with alum or mock PBS; mice were then boosted with the same regimen 22 d later and challenged 21 d thereafter. For all groups, as per protocol, animals were monitored daily for clinical signs of disease (hunching, ruffled fur and reduced activity) for the duration of the experiment. Weight loss was monitored daily for the first 7 d, after which weight monitoring continued until the animals recovered to their initial starting weight or displayed weight gain continuously for 3 d. All mice that lost greater than 20% of their starting body weight were ground-fed and further monitored multiple times per day as long as they were under the 20% cutoff. Mice that lost greater than 30% of their starting body weight were immediately sacrificed as per protocol. Any mouse deemed to be moribund or unlikely to recover was also humanely sacrificed at the discretion of the researcher. Euthanasia was performed using an isoflurane overdose and death was confirmed by cervical dislocation. All mouse studies were performed at the University of North Carolina (Animal Welfare Assurance #A3410-01) using protocols approved by the UNC Institutional Animal Care and Use Committee (IACUC).
Histological analysis.
The left lung was removed and submerged in 10% buffered formalin (Fisher) without inflation for 1 week. Tissues were embedded in paraffin and 5-μm sections were prepared by the UNC Lineberger Comprehensive Cancer Center histopathology core facility. To determine the extent of antigen staining, sections were stained for viral antigen using a commercially available polyclonal SARS-CoV anti-nucleocapsid antibody (Imgenex) and scored in a blinded manner by for staining of the airway and parenchyma as previously described20. Images were captured using an Olympus BX41 microscope with an Olympus DP71 camera.
Virus neutralization assays.
Plaque reduction neutralization titer assays were performed with previously characterized antibodies against SARS-CoV, as previously described11,12,13. Briefly, neutralizing antibodies or serum was serially diluted twofold and incubated with 100 p.f.u. of the different infectious clone SARS-CoV strains for 1 h at 37 °C. The virus and antibodies were then added to a 6-well plate with 5 × 105 Vero E6 cells/well with multiple replicates (n ≥ 2). After a 1-h incubation at 37 °C, cells were overlaid with 3 ml of 0.8% agarose in medium. Plates were incubated for 2 d at 37 °C, stained with neutral red for 3 h and plaques were counted. The percentage of plaque reduction was calculated as (1 − (no. of plaques with antibody/no. of plaques without antibody)) × 100.
Statistical analysis.
All experiments were conducted contrasting two experimental groups (either two viruses, or vaccinated and unvaccinated cohorts). Therefore, significant differences in viral titer and histology scoring were determined by a two-tailed Student’s t-test at individual time points. Data was normally distributed in each group being compared and had similar variance.
Biosafety and biosecurity.
Reported studies were initiated after the University of North Carolina Institutional Biosafety Committee approved the experimental protocol (Project Title: Generating infectious clones of bat SARS-like CoVs; Lab Safety Plan ID: 20145741; Schedule G ID: 12279). These studies were initiated before the US Government Deliberative Process Research Funding Pause on Selected Gain-of-Function Research Involving Influenza, MERS and SARS Viruses (http://www.phe.gov/s3/dualuse/Documents/gain-of-function.pdf). This paper has been reviewed by the funding agency, the NIH. Continuation of these studies was requested, and this has been approved by the NIH.
SARS-CoV is a select agent. All work for these studies was performed with approved standard operating procedures (SOPs) and safety conditions for SARS-CoV, MERs-CoV and other related CoVs. Our institutional CoV BSL3 facilities have been designed to conform to the safety requirements that are recommended in the Biosafety in Microbiological and Biomedical Laboratories (BMBL), the US Department of Health and Human Services, the Public Health Service, the Centers for Disease Control (CDC) and the NIH. Laboratory safety plans were submitted to, and the facility has been approved for use by, the UNC Department of Environmental Health and Safety (EHS) and the CDC. Electronic card access is required for entry into the facility. All workers have been trained by EHS to safely use powered air purifying respirators (PAPRs), and appropriate work habits in a BSL3 facility and active medical surveillance plans are in place. Our CoV BSL3 facilities contain redundant fans, emergency power to fans and biological safety cabinets and freezers, and our facilities can accommodate SealSafe mouse racks. Materials classified as BSL3 agents consist of SARS-CoV, bat CoV precursor strains, MERS-CoV and mutants derived from these pathogens. Within the BSL3 facilities, experimentation with infectious virus is performed in a certified Class II Biosafety Cabinet (BSC). All members of the staff wear scrubs, Tyvek suits and aprons, PAPRs and shoe covers, and their hands are double-gloved. BSL3 users are subject to a medical surveillance plan monitored by the University Employee Occupational Health Clinic (UEOHC), which includes a yearly physical, annual influenza vaccination and mandatory reporting of any symptoms associated with CoV infection during periods when working in the BSL3. All BSL3 users are trained in exposure management and reporting protocols, are prepared to self-quarantine and have been trained for safe delivery to a local infectious disease management department in an emergency situation. All potential exposure events are reported and investigated by EHS and UEOHC, with reports filed to both the CDC and the NIH.
Research in this manuscript was supported by grants from the National Institute of Allergy & Infectious Disease and the National Institute of Aging of the US National Institutes of Health (NIH) under awards U19AI109761 (R.S.B.), U19AI107810 (R.S.B.), AI085524 (W.A.M.), F32AI102561 (V.D.M.) and K99AG049092 (V.D.M.), and by the National Natural Science Foundation of China awards 81290341 (Z.-L.S.) and 31470260 (X.-Y.G.), and by USAID-EPT-PREDICT funding from EcoHealth Alliance (Z.-L.S.). Human airway epithelial cultures were supported by the National Institute of Diabetes and Digestive and Kidney Disease of the NIH under award NIH DK065988 (S.H.R.). We also thank M.T. Ferris (Dept. of Genetics, University of North Carolina) for the reviewing of statistical approaches and C.T. Tseng (Dept. of Microbiology and Immunology, University of Texas Medical Branch) for providing Calu-3 cells. Experiments with the full-length and chimeric SHC014 recombinant viruses were initiated and performed before the GOF research funding pause and have since been reviewed and approved for continued study by the NIH. The content is solely the responsibility of the authors and does not necessarily represent the official views of the NIH.
V.D.M. designed, coordinated and performed experiments, completed analysis and wrote the manuscript. B.L.Y. designed the infectious clone and recovered chimeric viruses; S.A. completed neutralization assays; L.E.G. helped perform mouse experiments; T.S. and J.A.P. completed mouse experiments and plaque assays; X.-Y.G. performed pseudotyping experiments; K.D. generated structural figures and predictions; E.F.D. generated phylogenetic analysis; R.L.G. completed RNA analysis; S.H.R. provided primary HAE cultures; A.L. and W.A.M. provided critical monoclonal antibody reagents; and Z.-L.S. provided SHC014 spike sequences and plasmids. R.S.B. designed experiments and wrote manuscript.
1. Ge XY, et al. Isolation and characterization of a bat SARS-like coronavirus that uses the ACE2 receptor. Nature. 2013;503:535–538. doi: 10.1038/nature12711. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
2. Yount B, et al. Reverse genetics with a full-length infectious cDNA of severe acute respiratory syndrome coronavirus. Proc. Natl. Acad. Sci. USA. 2003;100:12995–13000. doi: 10.1073/pnas.1735582100. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
3. Becker MM, et al. Synthetic recombinant bat SARS-like coronavirus is infectious in cultured cells and in mice. Proc. Natl. Acad. Sci. USA. 2008;105:19944–19949. doi: 10.1073/pnas.0808116105. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
6. He B, et al. Identification of diverse alphacoronaviruses and genomic characterization of a novel severe acute respiratory syndrome–like coronavirus from bats in China. J. Virol. 2014;88:7070–7082. doi: 10.1128/JVI.00631-14. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
7. Li F. Receptor recognition and cross-species infections of SARS coronavirus. Antiviral Res. 2013;100:246–254. doi: 10.1016/j.antiviral.2013.08.014. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
8. Sheahan T, et al. Mechanisms of zoonotic severe acute respiratory syndrome coronavirus host range expansion in human airway epithelium. J. Virol. 2008;82:2274–2285. doi: 10.1128/JVI.02041-07. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
9. Yoshikawa T, et al. Dynamic innate immune responses of human bronchial epithelial cells to severe acute respiratory syndrome–associated coronavirus infection. PLoS ONE. 2010;5:e8729. doi: 10.1371/journal.pone.0008729. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
10. Qiu X, et al. Reversion of advanced Ebola virus disease in nonhuman primates with ZMapp. Nature. 2014;514:47–53. doi: 10.1038/nature13777. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
11. Sui J, et al. Broadening of neutralization activity to directly block a dominant antibody-driven SARS-coronavirus evolution pathway. PLoS Pathog. 2008;4:e1000197. doi: 10.1371/journal.ppat.1000197. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
12. Sui J, et al. Effects of human anti–spike protein receptor binding domain antibodies on severe acute respiratory syndrome coronavirus neutralization escape and fitness. J. Virol. 2014;88:13769–13780. doi: 10.1128/JVI.02232-14. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
13. Rockx B, et al. Escape from human monoclonal antibody neutralization affects in vitro and in vivo fitness of severe acute respiratory syndrome coronavirus. J. Infect. Dis. 2010;201:946–955. doi: 10.1086/651022. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
14. Spruth M, et al. A double-inactivated whole-virus candidate SARS coronavirus vaccine stimulates neutralizing and protective antibody responses. Vaccine. 2006;24:652–661. doi: 10.1016/j.vaccine.2005.08.055. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
15. Bolles M, et al. A double-inactivated severe acute respiratory syndrome coronavirus vaccine provides incomplete protection in mice and induces increased eosinophilic proinflammatory pulmonary response upon challenge. J. Virol. 2011;85:12201–12215. doi: 10.1128/JVI.06048-11. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
17. Deming D, et al. Vaccine efficacy in senescent mice challenged with recombinant SARS-CoV bearing epidemic and zoonotic spike variants. PLoS Med. 2006;3:e525. doi: 10.1371/journal.pmed.0030525. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
18. Graham RL, Donaldson EF, Baric RS. A decade after SARS: strategies for controlling emerging coronaviruses. Nat. Rev. Microbiol. 2013;11:836–848. doi: 10.1038/nrmicro3143. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
19. Graham RL, Baric RS. Recombination, reservoirs and the modular spike: mechanisms of coronavirus cross-species transmission. J. Virol. 2010;84:3134–3146. doi: 10.1128/JVI.01394-09. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
20. Agnihothram S, et al. A mouse model for betacoronavirus subgroup 2c using a bat coronavirus strain HKU5 variant. MBio. 2014;5:e00047–14. doi: 10.1128/mBio.00047-14. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
21. Relman DA. Metagenomics, infectious disease diagnostics and outbreak investigations: sequence first, ask questions later? J. Am. Med. Assoc. 2013;309:1531–1532. doi: 10.1001/jama.2013.3678. [PubMed] [CrossRef] [Google Scholar]
23. Frieman M, et al. Molecular determinants of severe acute respiratory syndrome coronavirus pathogenesis and virulence in young and aged mouse models of human disease. J. Virol. 2012;86:884–897. doi: 10.1128/JVI.05957-11. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
24. Ren W, et al. Difference in receptor usage between severe acute respiratory syndrome (SARS) coronavirus and SARS-like coronavirus of bat origin. J. Virol. 2008;82:1899–1907. doi: 10.1128/JVI.01085-07. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
25. Sims AC, et al. Release of severe acute respiratory syndrome coronavirus nuclear import block enhances host transcription in human lung cells. J. Virol. 2013;87:3885–3902. doi: 10.1128/JVI.02520-12. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
26. Fulcher ML, Gabriel S, Burns KA, Yankaskas JR, Randell SH. Well-differentiated human airway epithelial cell cultures. Methods Mol. Med. 2005;107:183–206. [PubMed] [Google Scholar]
27. Roberts Anjeanette, Deming Damon, Paddock Christopher D., Cheng Aaron, Yount Boyd, Vogel Leatrice, Herman Brian D., Sheahan Tim, Heise Mark, Genrich Gillian L., Zaki Sherif R., Baric Ralph, Subbarao Kanta. A Mouse-Adapted SARS-Coronavirus Causes Disease and Mortality in BALB/c Mice. PLoS Pathogens. 2007;3(1):e5. doi: 10.1371/journal.ppat.0030005. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Quote Dr. David Martin said Peter Daszak wrote in 2015 ‘they needed the media to hype up the need for coronavirus vaccines to sell them’. The very next year Dr Ralf Baric wrote in 2016: ‘..’the (WIV) virus was human ready’ a clear indication a weapon was being readied for human transmission. Seems to me J. Jyrkkanen ‘Collusion and Conspiracy to commit a crime for profit’.
These folks woked together with a large team under the funding of Fauci, the Government and even President Biden to craft a deadly population whacking killer bioweapon and even created technology to hide the fact. The NoSeeUm Technology of Baric.
(Left to right): Ralph Baric, Peter Daszak, Shi Zhengli.
A group of Chinese scientists lobbied to rename ‘SARS-CoV-2’ to ‘2019-nCoV’. In the correspondence, the scientists feared that the virus would become known as ‘Wuhan Coronavirus’ or ‘Wuhan Pneumonia’.
The truth behind the science around the origin of the SARS-CoV-2 lies with Ralph Baric of the University of North Carolina, Peter Daszak of EcoHealth Alliance and Prof Shi Zhengli of the Wuhan Institute of Virology (WIV).
According to the email obtained by the US Right To Know organization, on January 13, 2020, in an email to Peter Daszak at 6.50 pm, Ralph Baric, states:
“Hi Peter, I have to participate in an NIH call tomorrow at 10. I believe it’s a strategic meeting designed to help craft a NIH response plan to the WU-CoV. Hope things are going well. Looks like we found our highly variable SARS-like CoV! Ralph”
In reply to Ralph Baric’s email, Peter Daszak at 7.55 pm states:
“It sounds like we’re on the same call! And my exact thoughts are of the highly variable SARS-like CoV. I’ve told journalists about it, but it’s a complicated story for them to get across.”
On 13 January 2020, before China or the World Health Organisation (WHO) made any official statement on the nature of the coronavirus, both Ralph Baric and Peter Daszak in their emails appear to be confident that the coronavirus in China is a “highly variable SARS-like CoV”. Most importantly, Ralph Baric refers to the coronavirus as “Our highly variable SARS-like COV”, displaying a familiarity with the virus.
Reportedly, in 2013, the American virologist Ralph Baric approached Zhengli Shi at a meeting. Shi had detected the genome of a new virus, called SHC014, that was one of the two closest relatives to the original SARS virus, but her team had not been able to culture it in the laboratory. Baric had developed a way around that problem—a technique termed as “reverse genetics” in coronaviruses. Not only did it allow him to bring an actual virus to life from its genetic code, but he could mix and match parts of multiple viruses. He wanted to take the “spike” gene from SHC014 and move it into a genetic copy of the SARS virus he already had in his laboratory. The spike molecule is what lets a coronavirus open a cell and get inside it. The resulting chimera would demonstrate whether the spike of SHC014 would attach to human cells.
Baric asked Shi Zhengli if he could have the genetic data for SHC014. “She was gracious enough to send us those sequences almost immediately,” he told media. His team introduced the virus modified with that code into mice and into a petri dish of human airway cells. Sure enough, the chimera exhibited “robust replication” in the human cells—evidence that nature was full of coronaviruses ready to leap directly to people.
It is no surprise that a group of Chinese scientists lobbied through US Professor of the University of North Carolina Ralph Baric to rename «SARS-CoV-2» given by the Coronavirus Study Group (CSG) of the International Committee on Virus Taxonomy (ICTV) to “2019-nCoV”. In the correspondence, the Chinese scientists feared that the virus would become known as “Wuhan Coronavirus” or “Wuhan Pneumonia”. In an email dated 13/2/2020, Professor Shi Zhengli wrote to Ralph Baric. The subject of the email was “Virus Name”. The email had an attached document titled, “A unique and unified name for the novel coronavirus from Wuhan SJ clean”.
The email stated:
“Dear Ralph,
We heard that the 2019-nCoV was renamed as SARS-CoV-2. We had a fierce discussion among Chinese virologists. We have some comments on this name, I’m wondering if the CoV study group would consider a revision.
I attached the comments from me and my Chinese colleague.”
The document from Prof. Shi Zhengli to Prof. Ralph Baric states:
“A unique and unified name is needed for the novel coronavirus identified from Wuhan. An outbreak of unusual pneumonia of unknown cause in Wuhan, China, was first reported in December 2019. By 5 January 2020, Chinese scientists had quickly identified the causative agent a new type of coronavirus (CoV) belonging to the Betacoronaviruses genus of the Coronavirdae family that also includes severe acute respiratory syndrome (SARS)-CoV and the Middle East respiratory syndrome (MERS)- CoV.
On 12 January 2020, the World Health Organization (WHO) temporarily named the virus as 2019 novel coronavirus (2019-nCoV). On 30 January, WHO recommended naming the disease as “2019-nCoV acute respiratory disease”. On 8 February 2020, the China National Health Commission (CNHC) announced naming the disease as “Novel CoronavirusPneumonia” (NCP). On 11 February 2020, WHO renamed the disease “coronavirus disease2019” (COVID-19). On 7 February 2020, the Coronavirus Study Group (CSG) of the International Committee on Virus Taxonomy (ICTV) posted a manuscript at bioRxiv and suggested designating the novel coronavirus as “severe acute respiratory syndrome coronavirus 2(SA RS-CoV-2)” based on the phylogenetic analysis of related coronaviruses.
Zhengli further in her document to Ralph Baric says, “By 11th February 2020, the new coronavirus had caused more than 40,000 confirmed infections and more than 1000 deaths, mostly in mainland China, despite efforts by the Chinese government and its people to contain the spread of the virus in past weeks. lt goes without saying that the effects of the epidemic on all the aspects of Chinese life are devastating and, possibly, irreversible. Consequently, appropriately naming the virus and disease becomes a matter of importance to the Chinese people, in general, and virologists, in specific, and the issue has been fervently discussed and debated among scientists with the outcome, so far, as noted above. We fully agree that the new virus and SARS-CoV belong to the same virus species by classification. However, the consensus opinion of Chinese virologists is that none of the currently proposed names reflects the uniqueness and characteristics of the novel virus and that more consideration is needed for naming the virus. Based on the following reasons, we propose giving a unique and unified name to the new virus.”
Prof Shi Zhengli, also known as the “Batwoman”, continues to impress upon Ralph Baric on behalf of the group of Chinese scientists. She says, “All proposed names are either too generic, or too similar, to previously well-known viruses, or contain an Arabic number. This makes it hard to remember or recognize, leading to a tendency among the general population and scientists alike to use a shorthand term such as ‘Wuhan coronavirus’ or ‘Wuhan pneumonia’. This has, in fact. been the case since it was named as 2019-nCoV. This practice would, however, stigmatize and insult the people in Wuhan, who are still suffering from the outbreak.”
The document sent by Prof Zhengli to Ralph Baric, further states, “The new virus is still evolving, and it is still too early to predict the outcome of the current outbreak. However, it is already clear that the infection of the new virus has diverse symptoms, from asymptomatic infection to severe pneumonia and even death. It has less case-fatality rate and higher transmissibility than SARS-CoV, indicating its clear difference from SARS-CoV. Again. therefore, it is not appropriate to designate the new virus as SARS-CoV-2 before we know more properties of the virus.”
Baric, Daszak and Zhengli were working together on the gain-of-function research. Scientists have posited that SARS-CoV-2 may be a product of WIV’s experiments on an unpublished bat coronavirus that is more closely related to SARS-CoV-2 than RaTG13. “First, SARS-CoV-2 may have evolved in bats, which are known reservoirs of immense coronavirus diversity, and then spread directly, or indirectly via an intermediate host, to humans through natural mechanisms. The degree of anticipated but undiscovered natural diversity clearly lends support to this scenario, as well as support to other scenarios. Second, SARS-CoV-2 or a recent ancestor virus may have been collected by humans from a bat or other animal and then brought to a laboratory where it was stored knowingly or unknowingly, propagated and perhaps manipulated genetically to understand its biological properties and then released accidentally.“
Wuhan Institute of Virology authorities shut down outside access to its virus database in September 2019, thereby making it difficult to verify that “The Wuhan lab has many bat samples not yet worked out or results published. There are some concerns that some of their samples may not have been handled properly and leaked out of the lab.”
Savio Rodrigues is the founder and editor-in-chief of Goa Chronicle.
Lab-made coronavirus related to SARS can infect human cells.
An experiment that created a hybrid version of a bat coronavirus — one related to the virus that causes SARS (severe acute respiratory syndrome) — has triggered renewed debate over whether engineering lab variants of viruses with possible pandemic potential is worth the risks.
In an article published in Nature Medicine1 on 9 November, scientists investigated a virus called SHC014, which is found in horseshoe bats in China. The researchers created a chimaeric virus, made up of a surface protein of SHC014 and the backbone of a SARS virus that had been adapted to grow in mice and to mimic human disease. The chimaera infected human airway cells — proving that the surface protein of SHC014 has the necessary structure to bind to a key receptor on the cells and to infect them. It also caused disease in mice, but did not kill them.
Although almost all coronaviruses isolated from bats have not been able to bind to the key human receptor, SHC014 is not the first that can do so. In 2013, researchers reported this ability for the first time in a different coronavirus isolated from the same bat population2.
The findings reinforce suspicions that bat coronaviruses capable of directly infecting humans (rather than first needing to evolve in an intermediate animal host) may be more common than previously thought, the researchers say.
But other virologists question whether the information gleaned from the experiment justifies the potential risk. Although the extent of any risk is difficult to assess, Simon Wain-Hobson, a virologist at the Pasteur Institute in Paris, points out that the researchers have created a novel virus that “grows remarkably well” in human cells. “If the virus escaped, nobody could predict the trajectory,” he says.
Creation of a chimaera
The argument is essentially a rerun of the debate over whether to allow lab research that increases the virulence, ease of spread or host range of dangerous pathogens — what is known as ‘gain-of-function’ research. In October 2014, the US government imposed a moratorium on federal funding of such research on the viruses that cause SARS, influenza and MERS (Middle East respiratory syndrome, a deadly disease caused by a virus that sporadically jumps from camels to people).
The latest study was already under way before the US moratorium began, and the US National Institutes of Health (NIH) allowed it to proceed while it was under review by the agency, says Ralph Baric, an infectious-disease researcher at the University of North Carolina at Chapel Hill, a co-author of the study. The NIH eventually concluded that the work was not so risky as to fall under the moratorium, he says.
But Wain-Hobson disapproves of the study because, he says, it provides little benefit, and reveals little about the risk that the wild SHC014 virus in bats poses to humans.
Other experiments in the study show that the virus in wild bats would need to evolve to pose any threat to humans — a change that may never happen, although it cannot be ruled out. Baric and his team reconstructed the wild virus from its genome sequence and found that it grew poorly in human cell cultures and caused no significant disease in mice.
“The only impact of this work is the creation, in a lab, of a new, non-natural risk,” agrees Richard Ebright, a molecular biologist and biodefence expert at Rutgers University in Piscataway, New Jersey. Both Ebright and Wain-Hobson are long-standing critics of gain-of-function research.
In their paper, the study authors also concede that funders may think twice about allowing such experiments in the future. “Scientific review panels may deem similar studies building chimeric viruses based on circulating strains too risky to pursue,” they write, adding that discussion is needed as to “whether these types of chimeric virus studies warrant further investigation versus the inherent risks involved”.
Useful research
But Baric and others say the research did have benefits. The study findings “move this virus from a candidate emerging pathogen to a clear and present danger”, says Peter Daszak, who co-authored the 2013 paper. Daszak is president of the EcoHealth Alliance, an international network of scientists, headquartered in New York City, that samples viruses from animals and people in emerging-diseases hotspots across the globe.
Studies testing hybrid viruses in human cell culture and animal models are limited in what they can say about the threat posed by a wild virus, Daszak agrees. But he argues that they can help indicate which pathogens should be prioritized for further research attention.
Without the experiments, says Baric, the SHC014 virus would still be seen as not a threat. Previously, scientists had believed, on the basis of molecular modelling and other studies, that it should not be able to infect human cells. The latest work shows that the virus has already overcome critical barriers, such as being able to latch onto human receptors and efficiently infect human airway cells, he says. “I don’t think you can ignore that.” He plans to do further studies with the virus in non-human primates, which may yield data more relevant to humans.
Quote Dr. David Martin said Peter Daszak wrote in 2015 ‘they needed the media to hype up the need for coronavirus vaccines to sell them’. The very next year Dr Ralf Baric wrote in 2016: ‘..’the (WIV) virus was human ready’ a clear indication a weapon was being readied for human transmission. Seems to me J. Jyrkkanen ‘Collusion and Conspiracy to commit a crime for profit’.
: “They took the money that Cheney gave them [from the Patriot Act], $2.2 billion, and they funneled it through NIH, and it all went through Anthony Fauci. So beginning in 2002, Anthony Fauci got a 68% raise from the Pentagon for doing bioweapons development, and he got a raise of billions of dollars a year, and then he started doing all of this gain-of-function. In 2014, three of those bugs escaped in high-profile escapes from different labs in the US. Congress held hearings on it. Everybody was angry, and 300 top scientists sent letters to Obama saying you got to shut down Fauci because he is going to create a pandemic. So, Obama ordered a moratorium, and at that time, Fauci had eighteen different gain-of-function experiments he was doing around the US. He instead moved his stuff offshore to Wuhan, where he could do it out of sight of these 300 scientists and nosy White House officials who were trying to shut him down. And he continued to do it with the same people he was funding here, Ralph Baric and Peter Dazak, and they moved their operation to the Wuhan lab.”
the CIA, DOD, and Tony Fauci taught Chinese military scientists how to build weapons of mass destruction. Then Bill Gates, a former CIA director, and China’s CDC director collaborated on how to censor a lab leak at Event 201 in Oct. 2019:
“Fauci funded the study that taught the Chinese military scientists, everything in China is dual-use, that lab is a military lab, and he taught them cutting-edge technology for building weapons of mass destruction.
In other words, the study for how to create the clones and how to create a spike protein that could attach to a human lung and transplant it onto a coronavirus.
He also funded through Ralph Baric at the University of North Carolina a technique called seamless ligation which is a technique for hiding human tampering on that virus after you’ve done it. Fauci gave Baric $212 million, and Baric developed a technique for hiding the human tampering; (Pic)
Quote Dr. David Martin said Peter Daszak wrote in 2015 ‘they needed the media to hype up the need for coronavirus vaccines to sell them’. The very next year Dr Ralf Baric wrote in 2016: ‘..’the (WIV) virus was human ready’ a clear indication a weapon was being readied for human transmission. Seems to me J. Jyrkkanen ‘Collusion and Conspiracy to commit a crime for profit’.
Baric taught that to Shi Zhengli (BELOW), the Chinese bat lady. Fauci says we were doing this for vaccine development and countermeasures, but there is no justification in the world for funding somebody to create seamless ligation; in fact, it is the inverse of what you would do if your interest was public health. If your interest was bioweapons creation, and he was the czar of bioweapons since 2002, that’s what you would do. USAID gave ten times what Fauci gave. The DOD was there. Why were they in there teaching Chinese scientists how to build weapons of mass destruction?
USAID is a CIA front group. Eco-health Alliance is a CIA front group. The CIA modeled this outbreak in 2019 twice, the second time at Event 201. Who was at Event 201? Avril Haines co-hosted it with Bill Gates, and the head of the Chinese CDC, George Gao, was there. The virus was already circulating in Wuhan, nobody knew it, but George Gao had to know it; he was the head of the Chinese CDC and their number one expert on coronaviruses. He comes to New York in October of 2019 and sits downs with Avril Haines, the former director of the CIA, today the Director of National Intelligence, the top spy in the country, and they do a four-part simulation, and the fourth part is George Gao and Avril Haines talking about how do we get social media to censor people if they say this is from a lab leak.”
.@RobertKennedyJr tells @jimmy_dore the CIA, DOD, and Tony Fauci taught Chinese military scientists how to build weapons of mass destruction. Then Bill Gates, a former CIA director, and China's CDC director collaborated on how to censor a lab leak at Event 201 in Oct. 2019:… pic.twitter.com/qYANxt4ZT2




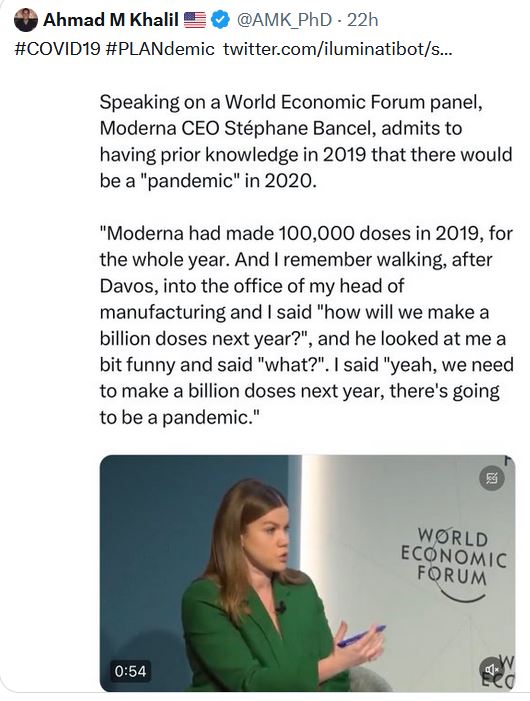











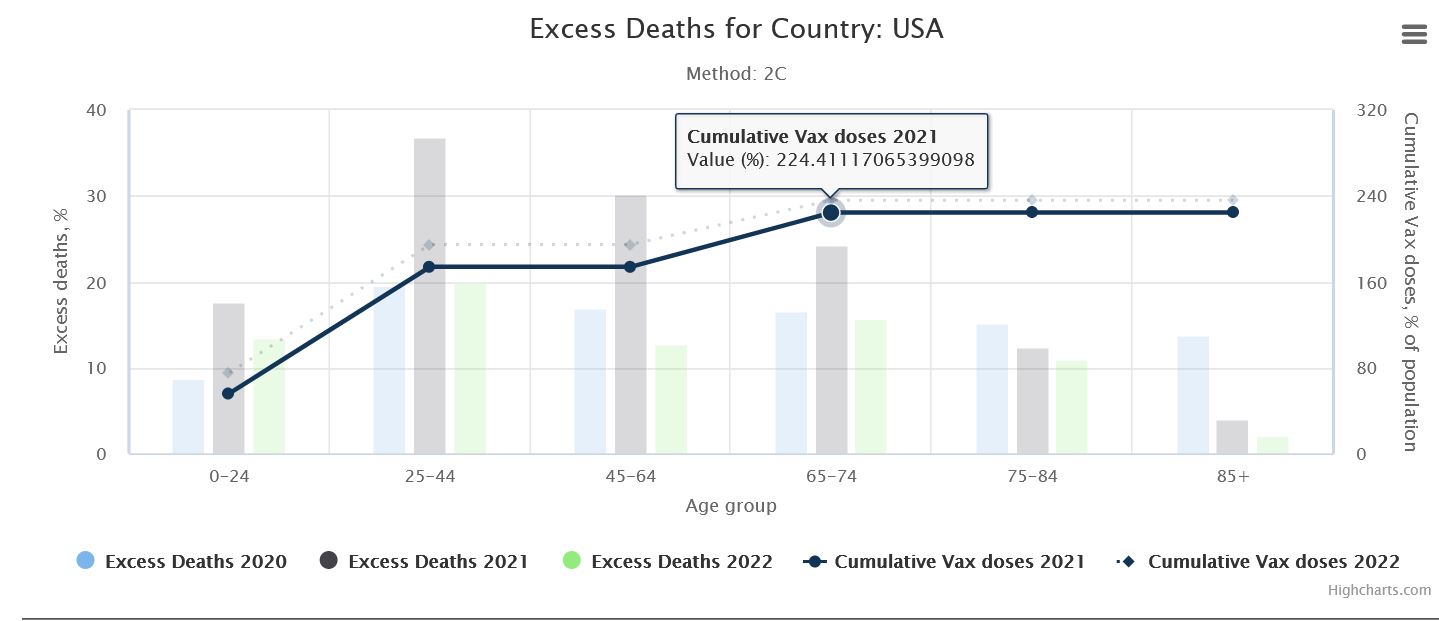
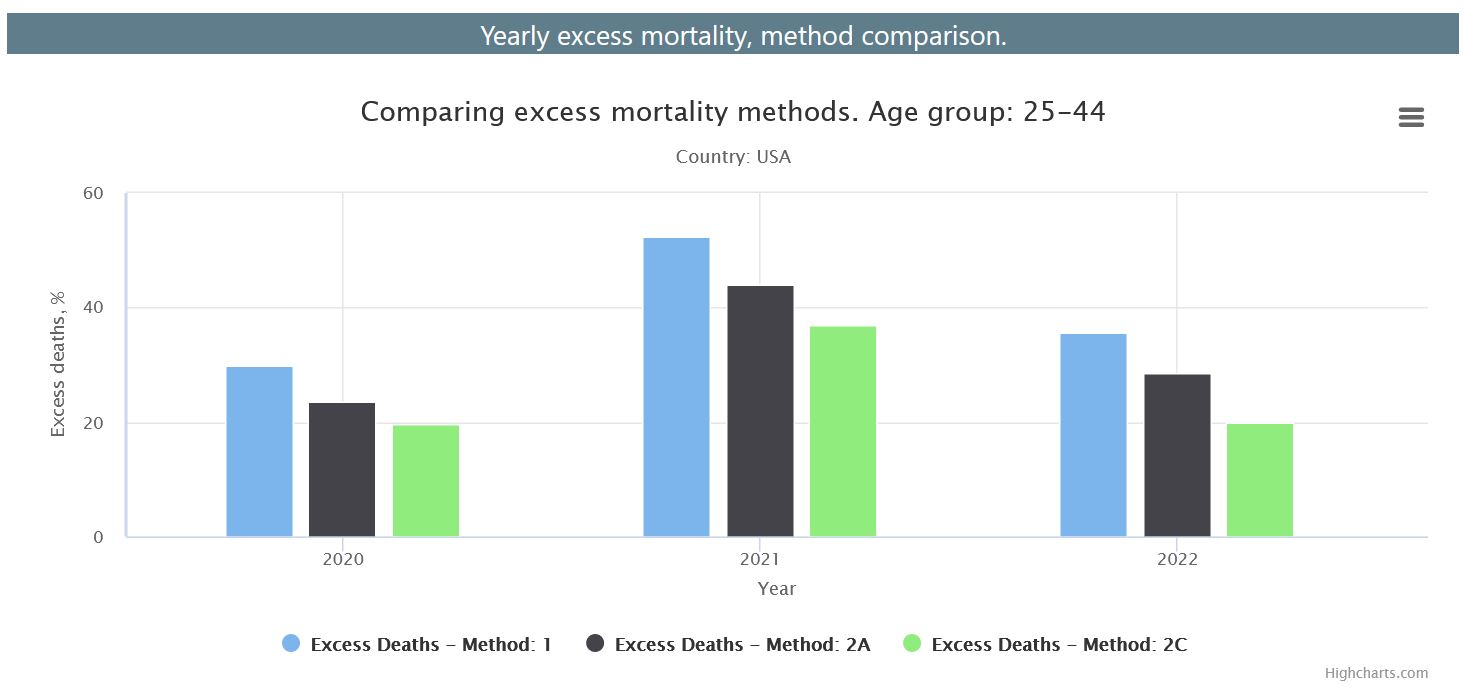













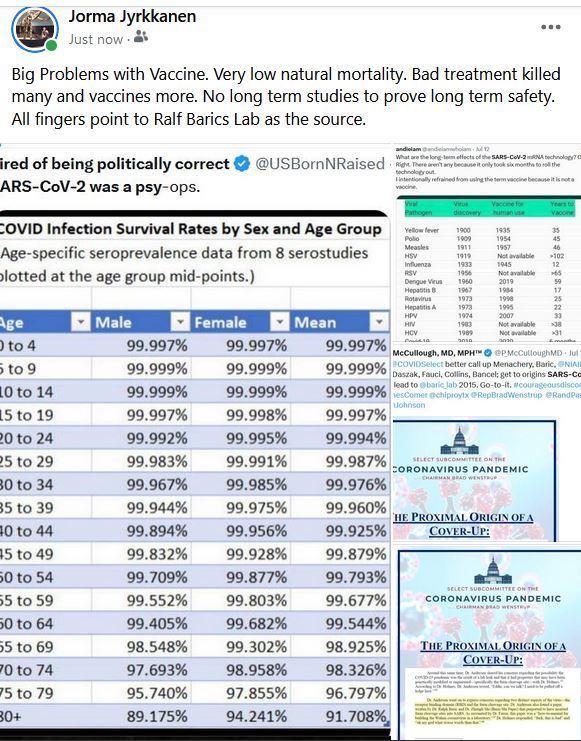



{kind=link}
{kind=link}
{kind=link}
{kind=link}